{kind=link}
Artículos científicos
Comportamiento productivo y niveles de ácidos grasos en la canal de corderos suplementados con Saccharomyces cerevisiae
Animal performance and fatty acids levels in lambs carcass suplemented with Saccharomyces cerevisiae
1Adrian Gloria Trujillo, 1*David Hernández Sánchez, 1 Omar Hernández Mendo, 1 María Magdalena Crosby Galván, 2 René Pinto Ruiz, 3 Edgar Meraz Romero, 1 J Efrén Ramírez Bribiesca
1 Postgrado en Ganadería, Instituto de Recursos Genéticos y Productividad, Colegio de Postgraduados, Campus Motecillo, km 36.5, Carretera México-Texcoco, Montecillo, Municipio de Texcoco, Edo. de México.*sanchezd@colpos.mx
2 Facultad de Ciencias Agronómicas, Universidad Autónoma de Chiapas.
3 Facultad de Medicina Veterinaria y Zootecnia. UNAM.
Artículo científico recibido: 18 de febrero de 2014
Aceptado: 02 de mayo de 2014
RESUMEN
Los ácidos grasos (AG) saturados depositados en la canal de rumiantes representan un punto crítico en la alimentación humana por su relación con enfermedades cardiovasculares. Ante esto, Saccharomyces cerevisiae (Sc) representa una alternativa para modificar el perfil de AG en la carne debido a sus elevados niveles de AG insaturados. Con este antecedente, se realizó un experimento utilizando 30 corderos comerciales en crecimiento (20 ± 0.9 kg PV), distribuidos en tres grupos de 10 animales cada uno, y cada grupo fue asignado a uno de tres tratamientos evaluados: 0, 3 y 5g de Sc animal-1 d-1, respectivamente, con el objetivo de determinar el comportamiento productivo del animal, niveles plasmáticos de colesterol y AG depositados en la canal. Se utilizó un diseño completamente al azar, los datos se analizaron usando PROC GLM, considerando diez repeticiones por tratamiento, y las medias de tratamientos se compararon con la prueba de Tukey. La adición de 3g animal-1 d-1 de Sc disminuyó (p < 0.05) parcialmente el consumo de MS y conversión alimenticia en los periodos evaluados, sin afectar (P > 0.05) la ganancia diaria de peso. Tampoco afectó (p > 0.05) el área del músculo Longissimus dorsi, espesor de grasa dorsal y colesterol total en sangre. Al incrementar la dosis de Sc, disminuyó (p < 0.05) el nivel de ácido mirístico, palmítico, oleico y linoleico. Se no afecta el comportamiento productivo del animal, tampoco mejora el perfil de AG insaturados en la canal; sin embargo, disminuye las concentraciones de AG saturados.
Palabras clave: Saccharomyces cerevisiae, corderos, oleico, canal, grasa dorsal.
ABSTRACT
Saturated fatty acids (FA) in the carcass of ruminants represent a critical topic in human alimentation given by their relationship with cardiovascular diseases. In view of this, Saccharomyces cerevisiae (Sc) represents an alternative to modify the meat FA profile by increasing the ratio of deposited unsaturated FA, because of its high levels of unsaturated fatty acids. An experiment was carried out using 30 commercial growing lambs (20 ± 0.9 kg LW), distributed into three groups with 10 animals each, which was placed into one of three evaluated treatments: 0, 3, and 5g Sc animal-1 d-1, in order to determine the productive performance of the animals, plasmatic cholesterol levels, and FA deposited in the carcass. A totally random design was used. Data were analyzed using the PROC GLM, considering ten repetitions per treatment. Mean comparison of the treatments was done through the Tukey test. The results show that adding 3g animal-1 d-1 Sc partially decreased (p < 0.05) DM intake and food conversion in the evaluated periods without affecting (P > 0.05) daily weight gain. It also did not affect (p > 0.05) muscle area of the Longissimus dorsi, fat thickness and total cholesterol in the blood. As the Sc dose increased, the levels of myristic, palmitic, oleic, and linoleic acids decreased (p < 0.05). Sc doesnt affect the animal performance, neither improvement the carcass unsaturated FA profile, however, saturated FA concentrations decreases.
Key words: Saccharomyces cerevisiae, lambs, oleic, carcass, fat thickness.
INTRODUCCIÓN
Saccharomyces cerevisiae (Sc) ha sido ampliamente evaluada como probiótico en estudios relacionados con la alimentación animal, tanto en rumiantes como en no rumiantes. Los estudios desarrollados en rumiantes contemplan evaluaciones relacionadas con la fermentación ruminal y su efecto en la respuesta animal (Sales 2011). Sin embargo, su utilización ha derivado en resultados variables (Duarte et al. 2012), encontrándose incrementos en las concentraciones de ácidos grasos volátiles después de suplementar Sc en la dieta, sugiriendo una mejora en la eficiencia de la fermentación ruminal y un mayor rendimiento debido a un incremento en la digestión de la fibra y en el consumo de materia seca (Zaworski et al. 2014). En contraste, en otras investigaciones el consumo y el pH ruminal no fueron afectados por el uso de la levadura (Allen y Ying 2012). Desafortunadamente, son pocos los reportes en la literatura que relacionan el efecto de Sc con los parámetros de calidad en la canal. Titi et al. (2008), reportaron incrementos en los niveles de grasa dorsal de corderos suplementados con Sc; sin embargo, no se menciona el efecto de la levadura sobre el perfil de la grasa depositada en la canal.
El cultivo de Sc se caracteriza por un perfil lipídico constituido de entre un 75.42 a 80.25 % de ácidos grasos insaturados (palmitoleico y oleico) (Tronchoni et al. 2012), lo que sugiere una fuente alterna de éstos ácidos grasos en el tejido adiposo del animal y de alguna forma podría disminuir los niveles de colesterol total, lipoproteínas de baja densidad y triglicéridos en el plasma sanguíneo del consumidor (Gnoni et al. 2010), debido a un menor consumo de ácidos grasos saturados en la carne, asociados con la presencia de enfermedades crónicas y cardiacas (Teira et al. 2006, Weingärtner et al. 2011). Por lo anterior, el objetivo del presente estudio fue determinar el comportamiento productivo de corderos en crecimiento, el perfil de ácidos grasos depositados en la canal y los niveles circulantes de colesterol, cuando se suplementa con niveles crecientes de Sc.
MATERIALES Y MÉTODOS
La presente investigación se realizó durante los meses de diciembre de 2010 y febrero de 2011, en la granja experimental del Colegio de Postgraduados, en Montecillo, Texcoco, Estado de México. Se utilizaron 30 corderos criollos, con peso vivo inicial promedio de 20 ± 0.9 kg. Se formaron tres grupos homogéneos, con diez animales cada uno, y distribuidos aleatoriamente en jaulas individuales. Los corderos fueron desparasitados (Ivomec F®) y vitaminados (Vigantol B® y Vigantol ADE Fuerte®) al inicio del estudio y 7 d posteriores.
El estudio tuvo una duración de 60 d, previo periodo de adaptación de 10 d. Se evaluaron tres tratamientos (T) con 10 repeticiones, variando el nivel de Saccharomyces cerevisiae (Sc) Yea Sacc1026 (levadura viable con una concentración de 1 x 108 UFC g-1): TI = Testigo, dieta comercial; T2 = TI + 3g de Sc animal-1 d-1 y T3 = TI + 5g de Sc animal-1 d-1. Las dosis de Sc se establecieron con base a las recomendaciones de suplementación de este aditivo alimenticio para corderos en engorda (Sales 2011), considerando un perfil de ácidos grasos de 8.8 % de mirístico, 26.8 % de palmítico, 6.1 % de esteárico, 25.7 % de oleico y 10.1 % de linoleico determinado por cromatografía de gases, según Blagovic et al. (2001). Se utilizó una dieta comercial para corderos en crecimiento con una composición proximal según análisis de laboratorio de 91.74 % de materia seca (MS), 15.71 % de proteína total (PT), 35.47 % de fibra detergente neutro (FDN) y 2.8 Mcal EM kg-1 MS, con un perfil de ácidos grasos de 14.69, 1.63, 29.93 y 53.74 % de palmítico, esteárico, oleico y linoleico, respectivamente. El agua se ofreció ad libitum y la dieta dos veces por día (8:00 y 16:00 h) ofreciendo el 5 % del peso vivo.
Comportamiento productivo
Para calcular el consumo de materia seca (CMS) se pesó diariamente el alimento ofrecido y rechazado. Los borregos se pesaron en ayuno cada 15 d para calcular la ganancia diaria de peso (GDP), realizando cuatro pesajes a lo largo del experimento, que conformaron los periodos de evaluación. Los datos de CMS y GDP, se usaron para calcular la conversión alimenticia (CA).
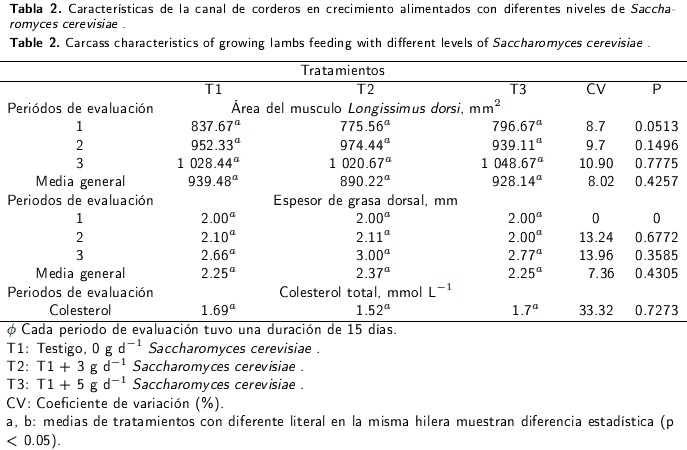
Características de la canal
Se evaluó el área de músculo Longissimus dorsi (LD) y el espesor de la grasa (EG) a la altura de la 12a y 13a costilla, con un equipo de ultrasonografía marca Medison Sonovet 600®, modelo SV-600 con un Transductor lineal de 7.5 Mhz. Las lecturas se obtuvieron en milímetros cuadrados para el área del ojo de la costilla y en milímetros para el espesor de grasa dorsal (Delfa et al. 1995).
Nivel de colesterol total en plasma sanguíneo
Se determinó el nivel de colesterol total en sangre. Para ello se tomaron muestras de sangre obtenidas de la vena yugular, utilizando tubos de recolección Vacutainer de 13 x 100 mm, y posteriormente fueron almacenadas en un contenedor con hielo, hasta ser procesadas. Las muestras se centrifugaron a 3200 x g durante 20 min en una centrifuga EBA 21 de Hettich, se extrajo el suero sanguíneo y fue congelado hasta su análisis. Se utilizó el método enzimático de punto final (Trinder 1969). La concentración de colesterol total (CT) (mg dL-1) se midió en un espectrofotómetro Modelo Varían Marca CARY 1E a una absorbancia de 505 nm.
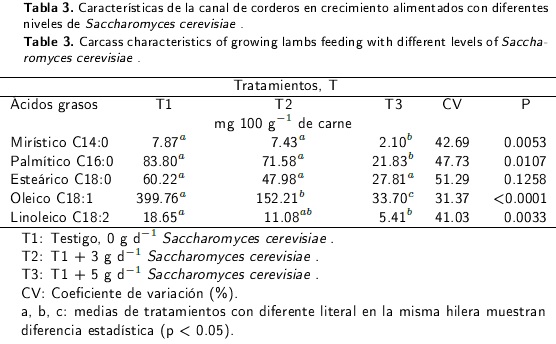
Perfil de ácidos grasos de Saccharomyces cerevisiae y en la canal
El perfil de ácidos grasos se determinó del músculo Longissimus dorsi, con base al método descrito por Folch et al. (1956). Las muestras se analizaron en un cromatógrafo de gases HP 6890 con un inyector HP 7683, equipado con una columna SPMR 2560, con dimensiones de 100 x 0.25 mm x 0.2 µm (película) a una presión de 199.94 kPa utilizando helio como gas acarreador (con un Make up de 18 mL min-1 a una presión de 199.94 kPa). La temperatura del inyector y detector fue de 250 y 260°C, respectivamente. La rampa de temperatura del horno se elevó de 110°C x 1 min a 240°C x 10 min. El contenido de ácidos grasos se expresó en mg 100 g carne-1.
Análisis estadístico
Se utilizó un diseño completamente al azar con tres tratamientos y 10 repeticiones. Los datos se analizaron mediante un procedimiento PROC GLM de SAS (SAS 2004). Para el análisis de las variables CMS, GDP y CA se utilizó el peso vivo inicial como covariable. La comparación múltiple de medias de tratamientos se realizó mediante la prueba de Tukey (α = 0.05).
RESULTADOS
Comportamiento productivo
Los resultados muestran que la adición de 3g d-1 de Sc en la dieta redujo (P < 0.05; Tabla 1) el CMS con respecto al grupo testigo durante el segundo periodo de evaluación. Así mismo, para la media general, el CMS fue de 1.25 vs 1.22 kg d-1 al suplementar 0 y 3g d-1 animal-1 de Sc. No obstante, la adición de 5 g d-1 de Sc (T3) no produjo cambios (p > 0.05) en esta variable. Para la GDP la adición de Sc no evidenció (p > 0.05) diferencias entre tratamientos. Mientras que la CA fue 21.7 % mejor (P < 0.05) en el T3 en comparación con el T2 en el tercer periodo de evaluación, sin embargo, ambos tratamientos no fueron diferentes al testigo.
Características de la canal
El área del músculo Longissimus dorsi (AML) no presentó cambios (p > 0.05; Tabla 2) entre tratamientos por efecto de la suplementación con Sc. Este comportamiento fue consistente con la media general. Así mismo, los valores de espesor de grasa (EG) tuvieron un comportamiento similar, sin diferencias (p > 0.05) entre tratamientos (Tabla 2).
Nivel de colesterol total en plasma sanguíneo
Los valores de colesterol total (CT) en plasma sanguíneo de los corderos alimentados con niveles crecientes de Sc no mostraron cambios (p > 0.05;Tabla 2) entre tratamientos, con promedios de 1.69, 1.52 y 1.7 mmol L-1 para T1, T2 y T3, respectivamente.
Características de la canal
El nivel de esteárico no fue diferente (p > 0.05; Tabla 3) entre tratamientos al adicionar Sc, pero si se obtuvo el nivel más bajo (p < 0.05) en el perfil del ácido mirístico, palmítico, oleico y linoleico depositados en la canal, a medida que se incrementaba el nivel de levadura a 5g animal-1 d-1.
DISCUSIÓN
Comportamiento productivo
El CMS en corderos suplementados con 3g d-1 de Sc fue menor (p < 0.05) en comparación con el grupo testigo. Este efecto es consistente con lo observado por Junniper et al. (2008), quienes reportaron que a medida que se incluye Sc en la dieta, el CMS se reduce al optimizarse el aprovechamiento de nutrientes (Fonty y Chaucheyras 2006), lo que permite al animal cubrir sus requerimientos nutricionales con menor cantidad de alimento (Di Francia et al. 2008, Moya et al. 2009). Sin embargo, esto contrasta con lo observado en el T3 (5g d-1 Sc) y con lo reportado por Haddad y Goussous (2005), Macedo et al. (2006) e Issakowicz et al. (2013) quienes indican que no hay efecto sobre esta variable al suplementar Sc en la dieta de corderos. Es por ello, que factores como el tipo de alimentación determinan el modo de acción de la levadura, explicando que en dietas a base de forraje, este aditivo genera mayor CMS, en comparación con dietas a base de concentrado (Kawas et al. 2007), ya que se incrementa la actividad bacteriana y la digestibilidad de la fibra (Zebeli et al. 2012, Hossain et al. 2012). La GDP no fue afectada (p > 0.05) por la suplementación con Sc y los valores obtenidos se mantuvieron dentro del intervalo reportado para corderos en engordas comerciales (200 a 300g d-1) (Souza et al. 2013). Al respecto se reporta que la respuesta en GDP por adicionar Sc en la dieta de borregos, solo es positiva cuando los niveles de proteína son menores a 15 % (NRC 2007, Tripathi y Karim 2011). Esta respuesta coincide con lo reportado por Titi et al. (2008), quienes al ofrecer una dieta con 16.8 % de proteína cruda obtuvieron valores similares entre el grupo testigo y el tratamiento con Sc (261 y 266g d-1, respectivamente). Congruente con estos reportes, en el presente estudio, la adición de Sc no afectó (p > 0.05) la GDP entre tratamientos, explicándose por el nivel de proteína de la dieta ofrecida (15.7 %). La falta de respuesta (p > 0.05) en la GDP y un CMS de 1.17 kg d-1 (p < 0.05), propiciaron, consecuentemente, mínima respuesta (p < 0.05) en la CA. Comportamiento similar reportaron Payandeh y Kafilzadeh (2007) y Sales (2011) quienes evaluaron Sc en una dieta para corderos.
Área del músculo Longissimus dorsi y espesor de grasa dorsal
La suplementación con Sc no generó cambios (p > 0.05) entre tratamientos al evaluar el AML y el EG. Consistente con esto, Kawas et al. (2007), Titi et al. (2008) y Rodrigues et al. (2013) reportaron una tendencia similar para corderos en finalización. Está documentado que factores como la edad, peso, raza y sexo del animal afectan el AML (Wilches et al. 2011), la cual disminuye cuando se incrementa la deposición de grasa dorsal, causada por la ingestión de dietas a base de grano (Seabrook et al. 2011). La falta de respuesta en estas variables, se puede explicar por la relación inversa que existe entre el AML y el EG, al disminuir la deposición de grasa cuando se incrementa la deposición de músculo (Rufino et al. 2013), debido a una mayor utilización de la energía y síntesis de proteína provocada por una mayor cantidad de proteína no degradada a nivel ruminal (Ponnampalam et al. 2001), la cual puede determinar la ausencia de diferencias significativas en el grado de engrasamiento, rendimiento y características de la canal, cuando los animales se alimentan bajo un mismo régimen alimenticio (Manso et al. 2009).
Colesterol total en plasma sanguíneo
De acuerdo a los resultados obtenidos en este trabajo, Sc (p > 0.05) no tiene efecto sobre las concentraciones plasmáticas de colesterol, ya que con niveles de 3 y 5 g animal-1 d-1, las concentraciones de este metabolito no cambiaron y fueron similares a los valores biológicos reportados para corderos en crecimiento alimentados con dietas a base de concentrado (1.61 a 4.98 mmol L-1) (Bodas et al. 2011); aun cuando los niveles de suplementación de Sc se incrementan a 15 y 30 g animal-1 d-1, los niveles plasmáticos de colesterol no son afectados (Pal et al. 2010). En este sentido, los resultados obtenidos en el presente estudio y los reportados en la literatura evidencian que la capacidad hipocolesterolemica de Sc es nula (Van Den Top et al. 1994, Beynen et al. 2000), señalando que el efecto principal de la levadura se limita a nivel ruminal (Sales 2011).
Perfil de ácidos grasos en la canal
La concentración de ácido oleico en la canal de corderos disminuyó (P < 0.05) a medida que se incrementaron los niveles de Sc en una dieta a base de concentrado. Esto contrasta con la concentración de ácido oleico reportada para corderos alimentados bajo condiciones intensivas, la cual se estima en los 669.7 a 860 mg 100 g-1 de carne (Nuernberg et al. 2008, Komprda et al. 2012). Está documentado que dietas a base de concentrado y altas en proteína explican que el ácido oleico represente hasta un 40 % del total de los ácidos grasos sintetizados en la canal de rumiantes (Duckett et al. 1993, Juárez et al. 2008) debido a las altas concentraciones de esteárico que llegan al duodeno (Loor et al. 2002) y son transformadas a ácido oleico por la actividad de la enzima delta 9 desaturasa (Castro et al. 2005), presente en bacterias ruminales, tejido adiposo e hígado (Valenzuela et al. 2009). Sin embargo, existen factores que inhiben esta enzima a diferentes niveles del metabolismo, como es el caso de la represión de los genes OLE1 en Sc (Cortés et al. 2009) y estearoil coenzima A desaturasa (SCD) en corderos (Daniel et al. 2004), al inhibir estos genes, disminuye la deposición de ácido palmítico, esteárico y oleico a nivel celular como ocurrió en el presente estudio, debido a que los principales sustratos para la síntesis de oleico usados por la enzima delta-9-desaturasa son el palmitoil-CoA y la estearoil-CoA (Wood et al. 2008). En este sentido, la presencia de los ácidos grasos de Sc pudo ejercer el efecto supresor del gen SCD y causar baja deposición de oleico como ocurrió en este estudio al suplementar con niveles crecientes de Sc.
CONCLUSIONES
Con base a las condiciones en que se desarrolló el estudio, se concluye que dosis crecientes de Sc en la dieta de corderos no afectan el comportamiento productivo del animal, tampoco mejoran el perfil de ácidos grasos insaturados en la canal; sin embargo, disminuye las concentraciones de ácidos grasos saturados, relacionados con la incidencia de enfermedades cardiovasculares. Por lo que se sugiere realizar mayor investigación que permita elucidar la dinámica y los efectos de Sc en la deposición de ácidos grasos en la canal.
AGRADECIMIENTOS
Los autores expresan sincero agradecimiento a la LPI 7, Inocuidad, Calidad de Alimentos y Bioseguridad, y a la LPI 11, Sistemas de Producción Agrícola, Pecuaria, Forestal, Acuícola y Pesquera del Colegio de Postgraduados, por el apoyo financiero para realizar esta investigación.
LITERATURA CITADA
Allen MS, Ying Y (2012) Effects of Saccharomyces cerevisiae fermentation product on ruminal starch digestion are dependent upon dry matter intake for lactating cows. Journal of Dairy Science 95: 6591-6605.
Beynen AC, Schonewille JT, Tersptra AHM (2000) Influence of amount and type of dietary fat on plasma cholesterol concentrations in goats. Small Ruminant Research 35: 141-147.
Blagovic B, Rupcic J, Mesaric M, Georgiú K, Vladimir M (2001) Lipid Composition of Brewer's Yeast. Food Technology Biotechnology 39(3): 175-181.
Bodas R, Prieto N, López CO, Giráldez FJ, Andrés S (2011) Naringin and vitamin E influence the oxidative stability and lipid profile of plasma in lambs fed fish oil. Research Veterinary Science 91: 98-102.
Castro T, Manso T, Mantecón AR, Guirao J, Jimeno V (2005) Fatty acid composition and carcass characteristics of growing lambs fed diets containing palm oil supplements. Meat Science 69: 757-764.
Cortés RC, Villagómez ME, Avalos SM, Molina AS (2009) Influencia de la peroxidación de lípidos sobre el daño oxidativo mitocondrial y la integridad de Saccharomyces cerevisiae. Information Tecnology 20(2): 71-81.
Daniel ZCTR, Wynn RJA, Salter M, Buttery PJ (2004) Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: The role of stearoyl-CoA desaturase. Journal of Animal Science 82: 747-758.
Delfa R, Teixeirab A, Gonzalez C, Blasco I (1995) Ultrasonic estimates of fat thickness and Longissimus dorsi muscle depth for predicting carcass composition of live Aragón lambs. Small Ruminant Research 16: 159-164.
Di Francia A, Masucci F, De Rosa G, Varricchio ML, Proto V (2008) Effects of Aspergillus oryzae extract and a Saccharomyces cerevisiae fermentation product on intake, body weight gain and digestibility in buffalo calves. Animal Feed Science and Technology 140: 67-77.
Duarte KMR, Gomes LH, Sampaio ACK, Issakowicz J, Rocha F, Granato TP, Terra SR (2012) Saccharomyces cerevisiae used as pobiotic: Strains characterization and cell viability. Journal of Agriculture and Veterinary Science 1(2): 17-19.
Duckett SK, Wagner DG, Yates LD, Dolezal HG, May SG (1993) Effects of time on feed on beef nutrient composition. Journal Animal Science 71: 2079-2088.
Folch J, Lees M, Sloane SGH (1957) A simple method for the isolation and purification of total lipids from animal tissues. Journal of Biology Chemistry 226: 497-509.
Fonty G, Chaucheyras FD (2006) Effects and modes of action of live yeasts in the rumen. Biología 61: 741-750.
Gnoni GV, Natali F, Geelen MJH, Siculella L (2010) Oleic Acid as an Inhibitor of Fatty Acid and Cholesterol Synthesis. Olives and Olive Oil in Health and Disease Prevention Chapter 152: 1365 - 1373.
Haddad SG, Goussous SN (2005) Effect of yeast culture supplementation on nutrient intake, digestibility and growth performance of Awassi lambs. Animal Feed Science and Technology 118: 343-348.
Hossain SA, Parnerkar S, Haque N, Gupta RS, Kumar D, Tyagi AK (2012). Influence of dietary supplementation of live yeast (Saccharomyces cerevisiae ) on nutrient utilization, ruminal and biochemical profiles of Kankrej calves. International Journal of Applied Animal Science 1(1): 30-38.
Issakowicz J, Bueno MS, Sampaio ACK, Duarte KMR (2013) Effect of concentrate level and live yeast (Saccharomyces cerevisiae ) supplementation on Texel lamb performance and carcass characteristics. Livestock Science 155: 44-52.
Juárez M, Horcada A, Alcalde MJ, Valera M, Mullen AM, Molina A (2008) Estimation of factors influencing fatty acid profiles in light lambs. Meat Science 79: 203-210.
Kawas JR, García CR, Garza CF, Fimbres DH, Olivares SE, Hernández VG, Luc CD (2007) Effects of sodium bicarbonate and yeast on productive performance and carcass characteristics of light - weight lambs fed finishing diets. Small Ruminant Research 67: 157-163.
Komprda T, Kuchtík J, Jarošová A, Dračková E, Zemánek L, Filipčík B (2012) Meat quality characteristics of lambs of three organically raised breeds. Meat Science 91: 499-505.
Loor JJ, Herbein JH, Jenkins TC (2002) Nutrient digestion, biohydrogenation, and fatty acid profiles in blood plasma and milk fat from lactating Holstein cows fed canola oil or canolamide. Animal Feed Science and Technology 97: 65-82.
Macedo R, Arredondo V, Beauregard J (2006) Influence of yeast culture on productive performance of intensively fattened Pelibuey lambs in Colima, México. Avances en Investigación Agropecuaria 10(3): 59-67.
Manso T, Bodas R, Castro T, Jimeno V, Mantecón AR (2009) Animal performance and fatty acid composition of lambs fed with different vegetable oils. Meat Science 83(3): 511-516.
Moya D, Calsamiglia S, Ferret A, Blanch M, Fandiño I, Castillejos L, Yoon I (2009) Effects of dietary changes and yeast culture (Saccharomyces cerevisiae ) on rumen microbial fermentation of Holstein heifers. Journal of Animal Science 87: 2874-2881.
NRC (2007) Nutrient Requirements of Small Ruminants. Sheep, goats, cervids, and new world camelids. Edition The National Academy Press, Washington, D.C. 362 p.
Nuernberg K, Fischer A, Nuernberg G, Ender K, Dannenberger D (2008) Meat quality and fatty acid composition of lipids in muscle and fatty tissue of Skudde lambs fed grass versus concentrate. Small Ruminant Research 74: 279-283.
Pal K, Paul SK, Biswas P, Patra AK, Bhunia T, Pakhira MC (2010) Responses of addition of yeast (Saccharomyces cerevisiae ) from rice distillers grains with solubles with or without trace minerals on the performance of Black Bengal kids. Small Ruminant Research 94: 45-52.
Payandeh S, Kafilzadeh F (2007) The effect of yeast (Saccharomyces cerevisiae ) on nutrient intake, digestibility and finishing performance of lambs fed a diet based on dried molasses sugar beet-pulp. Pakistan Journal Biology Science 10: 4426-4431.
Ponnampalam EN, Sinclair AJ, Egan AR, Blakeley SJ, Li D, Leury BJ (2001) Effect of dietary modification of muscle long-chain n?3 fatty acid on plasma insulin and lipid metabolites, carcass traits, and fat deposition in lambs. Journal of Animal Science 79: 895-903.
Rodrigues É, De Beni Arrigoni M, Mendonça ACR, Ludovico MC, Domingues MD, Parra FS, Mendes JA, Andrighetto C (2013) Performance, carcass characteristics and gain cost of feedlot cattle fed a high level of concentrate and different feed additives. Revista Brasileira de Zootecnia 42(1): 61-69.
Rufino LDA, Pereira OG, Ribeiro KG, Valadares FSC, Cavali J, Paulino PVR. (2013) Effect of substitution of soybean meal for inactive dry yeast on diet digestibility, lambs growth and meat quality. Small Ruminant Research 111: 56-62.
Sales J (2011) Effects of Saccharomyces cerevisiae supplementation on ruminal parameters, nutrient digestibility and growth in sheep: A meta-analysis. Small Ruminant Research 100: 19-29.
SAS (2004) SAS/STAT User's guide (Release 9.1). Cary, North Carolina, USA: SAS Institute.
Seabrook JL, Peel RK, Engle TE (2011) The effects of replacing dietary carbohydrate with calcium salts of fatty acids on finishing lamb feedlot performance, blood metabolites, muscle fatty acid composition, and carcass characteristics. Small Ruminant Research 95: 97-103.
Souza DA, Selaive VAB, Pereira ES, Osório JCS, Teixeira A (2013) Growth performance, feed efficiency and carcass characteristics of lambs produced from Dorper sheep crossed with Santa Inês or Brazilian Somali sheep. Small Ruminant Research 114: 51-55.
Teira G, Perlo F, Bonato P, Tisoco O (2006) Calidad de carnes bovinas. Aspectos nutritivos y organolépticos relacionados con sistemas de alimentación y sistemas de elaboración. Ciencia Docencia y Tecnología. 33: 173-193.
Titi HH, Dmour RO, Abdullah AY (2008) Growth performance and carcass characteristics of Awassi Lambs and Shami goat kids fed yeast culture in their finishing diet. Animal Feed Science and Technology 142: 33-43.
Trinder P (1969) Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Annals of Clinical Biochemistry 6: 24-27.
Tripathi MK, Karim SA (2011) Effect of yeast cultures supplementation on live weight change, rumen fermentation, ciliate protozoa population, microbial hydrolytic enzymes status and slaughtering performance of growing lamb. Livestock Science 135: 17-25.
Tronchoni J, Rozès N, Amparo Q, JM G (2012) Lipid composition of wine strains of Saccharomyces kudriavzevii and Saccharomyces cerevisiae grown at low temperature. International Journal of Food Microbiology 155: 191-198.
Valenzuela SAR, Montero LM, Juárez LFI (2009) Contenido de ácidos grasos y conjugados del ácido linoleico en carne de bovinos. Revista Electrónica de Veterinaria 10: 1-84.
Van Den Top AM, Wensing T, Beynen AC (1994) The influence of calcium palmitate and oleate feeding on hepatic lipid metabolism in dry goats. Journal of Animal Physiology and Animal Nutrition 72: 44-55.
Weingärtner O, Lütjohann D, Vanmierlo T, Müller S, Günther L, Herrmann W, Böhm M, Laufs U, Herrmann M (2011) Markers of enhanced cholesterol absorption are a strong predictor for cardiovascular diseases in patients without diabetes mellitus. Chemistry Physiology Lipids 164: 451-456.
Wilches D, Rovira J, Jaime I, Palacios C, Luruena MMA, Vivar QAM, Revilla I (2011) Evaluation of the effect of a maternal rearing system on the odour profile of meat from suckling lamb. Meat Science 88: 415-423.
Wood JD, Enser M, Fisher AV, Nute GR, Sheard PR, Richardson Rl, Hughes SI, Whittington FM (2008). Fat deposition, fatty acid composition and meat quality: A review. Meat Science 78: 343-358.
Zaworski EM, Shriver-Munsch CM, Fadden NA, Sanchez WK, Yoon I, Bobe G (2014) Effects of feeding various dosages of Saccharomyces cerevisiae fermentation product in transition dairy cows. Journal of Dairy Science 97(4): 3081-3098.
Zebeli Q, Aschenbach JR, Tafaj M, Boguhn J, Ametaj BN, Drochner W (2012) Invited review: Role of physically effective fiber and estimation of dietary fiber adequacy in high-producing dairy cattle. Journal of Dairy Science 95: 1041-1056.