Articulo Cientifico
INCREMENTO DE EMBRIONES SOMÁTICOS DE CACAO (Theobroma cacao L.) EN SISTEMA DE INMERSIÓN AUTOMATICO
Increase of embryos somatic of Cocoa (Theobroma cacao L.) in Immersion System Automatic
Jorge Luis Peña-López1, Alfonso Azpeitia-Mora les2*, Felipe Mirafuentes-Hernández2, Violeta Ruíz-Carrera, Luis Sáenz-Carbonell3
1 División Académica de Ciencias Biológicas. Universidad Juárez Autónoma de Tabasco. km 0.5, s/n, entronque a Bosques de Saloya, CP. 86150, Villahermosa, Tabasco, México
2 Laboratorio de Biología Molecular. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Huimanguillo, km 1, Carretera Huimanguillo-Cárdenas.
3 Centro de Investigación Científica de Yucatán, A.C. Chuburna de Hidalgo, Mérida, Yucatán. *Autor de correspondencia: azpeitia20032003@yahoo.com.mx
Artículo científico recibido el 23 de julio de 2015
Aceptado el 21 de octubre de 2015
RESUMEN
El cacao es un árbol tropical originario de América, su fruto se usa en la elaboración del chocolate y sus derivados. La propagación del cacao se realiza por la vía sexual o semillas, adicionalmente se propaga por enraizamiento de estacas e injertos. En la actualidad también se emplea la embriogénesis somática para la propagación clonal del cacao, la cual puede repetirse varias veces, lo que contribuye al incremento en la formación de embriones somáticos; por lo que el uso de un sistema de inmersión automático, puede incrementar la producción de embriones somáticos secundarios. Por lo anterior, el objetivo fue evaluar el sistema de inmersión automático Rita® en el rendimientode embriones somáticos secundarios de los 30 a los 180 d en dos ciclos de cultivo. Los resultados muestran que se generaron embriones somáticos en los explantes de fragmento de cotiledón y en embriones somáticos completos. Se obtuvo un segundo ciclo de la embriogénesis somática secundaria, en el que se obtuvieron 150 embriones somáticos por recipiente Rita® con promedio de 15 embriones por explante. El sistema de inmersión automático favoreció laproducción de embriones somáticos, su sincronización y disminuyó la oxidación del tejido.
Palabras clave: Cacao, Theobroma cacao L, embriogénesis somática secundaria, inmersión temporal
ABSTRACT
Cacao is a tropical tree native to America. Its fruit is used in the elaboration of chocolate and its derivatives. The propagation of cacao is done through sexual means or seeds; additionally, it propagates through theuse of cuttings and grafting. Currently, somatic embryogenesis is also employed for the clonal propagation of cacao, which can be repeated several times and contributes to the increase in the formation of somatic embryos. Therefore, theuse of a temporary immersion system can increase the production of secondary somatic embryos. Given the aforemen-tioned, the objective was to evalúate the Rita® temporary immersion system in the performance of secondary somaticembryos from 30 to 180 days in two cultivation cycles. The results showed that somatic embryos were generated incotyledon explants and in full somatic embryos. A second cycle was obtained from the secondary somatic embryogenesis where 150 somatic embryos were obtained per Rita® recipient with an average of 15 embryos per explant.The temporary immersion system favored the production of somatic embryos, its synchronization, and decreased theoxidation of the tissue.
Key words: Cacao, Theobroma cacao L., secondary somatic embryogenesis, temporal immersion
INTRODUCCION
El cacao es un árbol originario de América, su fruto es la base para la elaboración de chocolate y sus derivados (Borrone et al. 2007). Esta especie se cultiva en los trópicos húmedos y es fuente importante de ingresos en los países en desarrollo (Lanaud et al. 2009). La propagación del cacao se realiza generalmente por la vía sexual o semillas (Minyaka et al. 2008). Por lo que el 70 % de los árboles de cacao cultivados en el mundo provienen de semilla sin selección previa (Eskes 2000). Aunque también, se propaga por métodos de reproducción asexual ovegetativa, por enraizamiento de estacas e injerto (Salvador et al. 2005).
En la actualidad, el empleo de la embriogénesis somática (ES) para la propagación clonal de plantas de cacao es de gran importancia (Yang y Zhang 2010). Este es un proceso en el que una estructura bipolar parecida a un embrión cigótico se desarrolla a partir de un tejido somático sin conexión vascular con el tejido original (Zahid et al. 2013). Las células atraviesan por una serie de cambios morfológicos y bioquímicos que dan como resultado la noformación de embriones somáticos y la regeneración de nuevas plantas (Komamine et al. 2005). Los embriones somáticos (ESO) son parecidos a los embriones cigóticos y tienen también los mismos estados de desarrollo (Yang y Zhang 2010). Alternativamente, los ESO pueden ser utilizados como explantes para producir ESO adicionales a través de la embriogénesis somática secundaria (ESS) ns (Alejandro-Lázaro et al. 2015). La ESS puede ser repetida varias veces, por lo tanto, contribuye de forma significativa al incremento de la formación de embriones somáticos, especialmente en las plantas de baja capacidad embriogénica (Pérez-Núñez et al. 2006). La ESS también ha sido reportada para ayudar a retener la competencia embriogénica durante prolongados tiempos de cultivo en diferentes especies (Martinelli et al. 2001). Por otra parte, los embriones obtenidos a través de la ESS suelenser más desarrollados que los obtenidos por embriogénesis somática primaria (Pérez-Núñez et al. 2006). Desde las primeras observaciones de la embriogénesis somática, su potencial ha sido mostradoy caracterizado en un amplio número de plantas dicotiledóneas y monocotiledóneas (Mathieu et al. 2006). Diversos estudios para el desarrollo de protocolos para la propagación clonal de cacao por cultivode tejidos se han reportado (Alejandro-Lázaro et al. 2015), pero los resultados más relevantes se han obtenido por ES, con fuentes de explante de embriones cigóticos, estaminoides y pétalos, pero conbaja eficiencia y bajas tasas de conversión a plántula (Urrea et al. 2011).
En la actualidad, se tienen avances en el desarrollo de sistemas de cultivo de tejidos de ES de cacao, lo que puede contribuir en el mejoramiento de las variedades (Maximova et al. 2008). Sin embargo, los resultados indican que muchos genotipos de cacao son recalcitrantes (Minyaka et al. 2008), por lo que se ve afectada la ES (Tan y Furtek 2003). Actualmente, dos limitaciones principales en la ES de cacao son la eficacia de producción de embrión dependiente del genotipo y la falta de desarrollo de cotiledones en los embriones somáticos, lo que provoca bajo porcentaje de conversión de planta (Maximova et al. 2014).
Los métodos utilizados para la propagación in vitro de plantas, presentan ventajas sobre los métodos tradicionales, como mínima superficie necesaria para mantener una gran cantidad de plantas, obtención de plantas libres de hongos, bacterias y/o virus, y la posibilidad de producir plantas todo el año (Albarrán et al. 2005). Debido a estas características, existen medios para cultivos in vitro en estado semisólido y líquido (Watt 2012). Los medios semisólidos son aquellos a los cuales se les agrega un agente gelificante, son los más utilizados en el establecimiento de explantes (Ziv 2005). Mientras que el medio líquido y el empleo de un sistema de inmersión temporal son adecuados para la producción y calidad de embriones somáticos (Albarrán et al. 2005). La inmersión periódica delos explantes en el medio de cultivo, beneficia al intercambio gaseoso dentro del recipiente y facilitala absorción de nutrientes (García-Gonzáles et al. 2010). Lo que se refleja en una mayor tasa de multiplicación y en un mejor desarrollo de los explantes (Albarrán et al. 2005). Por lo anterior el objetivo del presente estudio fue evaluar el sistema de inmersion temporal Rita® en el rendimiento de embriones somaticos secundarios de los 30 a los 180 d en dos ciclos de cultivo
MATERIALES Y MÉTODOS
Colecta y desinfección de botones florales
Se colectaron botones florales del genotipo íse híbrido INIFAP 1 entre las ocho y 10 h, los cuales se depositaron en frascos de Gerber que contenían agua bidestilada estéril. Para luego trasladarlos al laboratorio donde se desinfectaron en una campana de flujo laminar, con una solución de hipoclorito de sodio al 1 % durante 20 min. Para luego realizar tres lavados con agua destilada estéril, para posteriormente cultivarse las bases de los pétalos de las flores en cajas Petri.
Inducción de embriogénesis somática primaria
Se utilizó el medio de cultivo denominado Primary Callus Growth (PCG) suplementado con 5 pg de Thidiazuron y 200 pg de 2,4-D. Los cultivos se mantuvieron en obscuridad por 14 d a temperatura de 26 ± 1 °C. Para luego transferir los explantes a un medio de crecimiento de callos secundarios (SCG) y continuar con la formación de callo embriogénico en ED4 (Driver y Kuniyuki 1984). Para luego transferir a un medio EDL de desarrollo de embriones somáticos para la regeneración de plantas en medio PR (Li et al. 1998). Durante toda la etapa el cultivo se mantuvo en obscuridad y a temperatura de 26 ± 1 °C, renovando el medio de cultivo cada 14 d.
Inducción de embriogénesis somática secundaría a partir de cotiledones
Se usaron fragmentos de cotiledones de 5 mm2 procedentes de embriones somáticos primarios del híbrido INIFAP 1, generados después de cuatro meses de cultivo. Los fragmentos se cultivaron en un medio de regeneración de plantas PR (Figura1) (Driver y Kuniyuki 1984). Suplementado con 30 gL-1 de sacarosa, 5 gL-1 de glucosa, 0.2 gL-1 de nitrato de potasio y 3 gL-1 de Gelrite comogelificante. El pH fue ajustado a 5.8 con hidróxidode sodiol N.
Primer ciclo de embriogénesis somática secundaria
Se emplearon explantes del embrión somático tipo cotiledónar y fragmentos de cotiledón de 5mm2. Los cuales se colocaron en un medio PR confitohormonas para regenerar las plantas, desprovisto de fitohormonas. No se realizaron subcultivos durante el ciclo de cultivo. El experimento se estableció bajo un diseño experimental completamente al azar en arreglo factorial 3x2 con cinco repeticiones. Las variables estudiadas fueron el númerode embriones somáticos formados y el porcentajede oxidación. Los resultados se analizaron con un análisis de varianza y comparación de medias deTukey (0.05). Los tratamientos evaluados se presentan en la Tabla 1.
Segundo ciclo de embriones somáticos secundarios
Un segundo experimento fue establecido para evaluar la capacidad de multiplicación de los embriones somáticos secundarios completos durante un segundo ciclo, partiendo de los embriones somáticos generados del experimento anterior. Solamente se utilizaron embriones somáticos procedentes del sistema Rita donde fueron cultivados. Los tratamientos en evaluación se presentan en la Tabla 1.
En este experimento se utilizó un solo tipo de explante: embrión somático tipo cotiledónar. El medio de cultivo utilizado fue PR (regeneración de plantas) desprovisto de fitohormonas. No se realizaron subcultivos a lo largo de 180 días de duración de este ciclo de cultivo.
El experimento se estableció bajo un diseño experimental completamente al azar con tres tratamientos (Tabla 1) con cinco repeticiones y cada repetición fue establecida con 10 explantes (embriones somáticos secundarios). Las variables en estudio fueron: número de embriones somáticos formados y porcentaje de oxidación de embriones somáticos. Los resultados fueron examinados por medio de un análisis de varianza y prueba de comparación de medias de Tukey (0.05).
Descripción del proceso histológico
El proceso histológico se realizó de acuerdo a Buffard-Morel et al.(1992) en muestras de fragmentos de cotiledón del sistema de inmersión automático a los 180 d. Se utilizó el estuche de Polysiences, Inc (USA) JB-4 a base de glicolmetacrilato. Las muestras se fijaron en paraformaldehído al 4 % en amortiguador de fosfatos (pH = 7.2) por 24 h bajo presión negativa, posteriormente se deshidrataron en las concentraciones de etanol 30 % por 30 min, 50 % por 45 min, 70 % por 45 min, 90 % por 60 min, 95 %por 60 min y 100 % por 60 min dos veces. Para la impregnación de los tejidos se utilizó mi desolución A (monómero) y 900 mg del catalizador C (Bemzoil peróxido) por 24 h. Por último, se realizó la inclusión de las muestras en 25 mL deuna solución A + C 25 mL, a la cual se le adicionó 1 mL de solución B (Polietilen glicol), para luego agregar al molde la solución conteniendo las muestras. A partir de los tejidos impregnados en resina, se obtuvieron cortes histológicos de 3 con un micrótomo Microm 325 (USA). La tinción se realizó con ácido periódico al 1 % por 5 min, para luego aplicar azul de metileno por 5 min, seguido de lavados con agua estéril. Posteriormente, se colocaron las muestras en una parrilla de calentamiento a 25 °C para su secado. Las secciones se observaron y fotografiaron con un microscopio Cari Zeiss (Alemania) con cámara digital acoplada.
RESULTADOS
Primer ciclo de embriogénesis somática secundaría
Después de los 30 d, todos los tratamientos fueron similares, mostrando escasa formación de embriones somáticos (ESO). Sin embargo, a los 120 d el tratamiento correspondiente a fragmentos de cotiledón fue estadísticamente superior al resto de los tratamientos con 26 ESO, mientras que con el tratamiento en recipiente Magenta sólo se obtuvieron ocho ESO. Al final del experimento, los cultivos en el sistema de inmersión temporal Rita® fueron estadisticamente superiores al testigo (Magenta®), en tanto que en el sitema Rita® se tuvieron 30 ESO generados con fragmentos de cotiledón y 29 ESO en cotiledones completos, produciéndose por explante en promedio tres ESO.
Mientras que en el recipiente Magenta® se obtuvieron cinco ESO de explantes de embriones somáticos completos y ocho ESO de fragmentos de cotiledón. Los resultados muestran que la producción de embriones fue mayor a los 180 d de cultivo en el sistema Rita®, mientras que en el recipiente Magenta la mayor producción de embriones se presentó a los 120 d de cultivo en fragmentos de cotiledón (Figura 1). En la Figura 2 se muestran los explantes utilizados y los embriones generados en el Sistema de Inmersion Temporal Rita®.
Porcentaje de oxidación
En los tratamientos, el porcentaje de oxidación incrementó con el tiempo, pero a los 180 d el sistema Rita® con embriones somaticos completos tuvo el menor porcentaje de oxidación con 45 %, mientras que el sistema Rita® con fragmentos de cotiledón presentó un 55 %, siendo estos tratamientos superiores estadísticamente a los tratamientos em medio semisolido en los recipientes Magentalos cuales tuvieron un 90 % de oxidación en embriones somáticos completos y 100 % en fragmentos de cotiledón (Figura 3).
Producción de embriones somáticos en un segundo ciclo
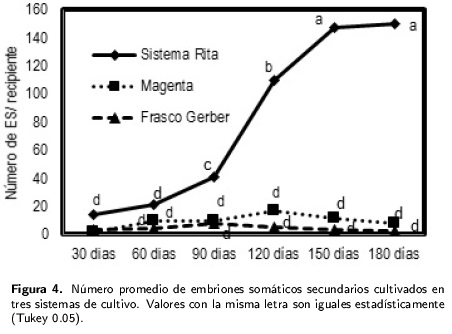
A los 30 d de cultivo se tuvieron embriones tipo globular, que se desarrollaron en el tejido superficial, apareciendo en mayor proporción en el sistema Rita® (Figura 4), con promedio de 0.84 ES a los 30 d. El mayor rendimiento de embriones somáticos secundarios se tuvo a los 180 d con promedio de 15 ES por recipiente. Este tratamiento superó estadísticamnete al testigo en frascos tipos Gerber® y a los recipientes Magenta®.
En el testigo, la respuesta a la formación de embriones somáticos fue de 0.31 ES a los 30 d, obteniéndose la máxima formación (1.7 ES) alos 90 d (Figura 4), con embriones de tipo globular que se observaron en pequeños grupos de color verde traslucidos. En el tratamiento Magenta® se obtuvo la menor cantidad de embriones somáticos formados a los 30 d con 0.16 ES en promedio, incrementándose hasta alcanzar su mayor promedio a los 120 d con 1.73 ES (Figura 4), con embriones de tipo cotiledónar en menor tiempo que el resto de los tratamientos. Estos embriones tuvieron una coloración amarillenta a los 90 d de cultivo.En la Figura 5 se muestran embriones somáticos globulares y un embrión somático tipo torpedo y sus cortes histológicos.
Porcentaje de oxidación
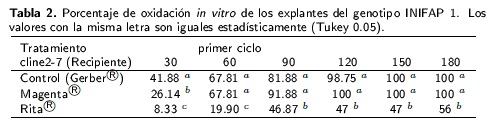
Todos los tratamientos presentaron diferentes porcentajes de oxidación a partir del primer mes decultivo. El tratamiento en sistema Rita® mostró el porcentaje más bajo de oxidación durante todo el experimento (Tabla 2). A los 180 d de cultivo, el tratamiento presentó en 56 % de oxidación de tejido siendo superior estadísticamente a los tratamientos en recipientes Gerber y Magenta®. La aparición de pequeños puntos cafés fue un indicador del comienzo oxidativo. Los explantes establecidosen el tratamiento testigo se oxidaron por completo a los 150 d, mientras que en el recipienta Magenta® la oxidación total se presentó a los 120 d.
DISCUSION
El sistema de inmersión automático fue desarrollado para mejorar la propagación in vitro, de cultivos de importancia comercial (Watt 2012). En la presente investigación, los resultados mostraron que el uso de medio liquido en el sistema Rita® favoreció la producción de embriones somáticos, por lo que la baja producción de embriones somáticos secundarios en medios semisólidos está asociada al agotamiento de los nutrimentos (Ziv 2005). Por lo que los medios líquidos en el sistema de inmersion automatico Rita® favorecen la producciónde embriones somáticos debido a que el explante entra en mayor contacto con el medio, lo que facilita la absorción de nutrimentos (García-Gonzáles 2010). Adicionalmente, este sistema ha demostrado ser positivo en la producción y calidad de embriones somáticos de café (Albarrán et al. 2005), resultados similares se observaron en el presente estudio, con embriones somáticos secundarios independientes sin fusiones o malformaciones. Se observó que la exclusión de fitohormonas favorece la multiplicación y la sincronización de los embriones de forma independiente sin fusiones. Estos resultados son similares a los reportados por Alejandro-Lázaro et al. (2015) quienes indican que generaron ESS en cacao en un medio semisólido sin fitohormonas. La formación de ESS se pudo repetir en un segundo ciclo de cultivo, con disminución del porcentaje de oxidación en el sistema de inmersión automático Rita®. Al respecto Da miaño et al. (2005) reportan que la disminución de la oxidación se debe a la renovación de la atmósfera dentro del recipiente, lo que permite reducir los niveles de contaminación y la hiperhidricidad de los tejidos. El uso de la ESS en cacao es una herramienta valiosa en la propagación de plantas de genotipos agronómicamente importantes (Maximova et al. 2008). Además de que los resultados obtenidos son superiores a lo reportado por Alejandro-Lázaro et al. (2015), incluso se logró la multiplicación de embriones somáticos en un segundo ciclo de cultivo, con estructuras como cotiledones, eje embrionario y haces vasculares. El potencial para usar embriones somáticos para la propagación a gran escala de genotipos élite de cacao puede representar un avance importante para la multiplicación de genotipos.
CONCLUSIONES
El uso de medio líquido en sistema Rita® favoreció la producción de embriones somáticos de cacao. Los embriones somáticos secundarios obtenidos no presentaron fusiones o malformaciones recurrentes. La cual fue posible repetirla en un segundo ciclo de cultivo, demostrando la totipotencia de los embriones somáticos completos usados como explantes, al igual que los fragmentos de cotiledón y sobre todo la repetitividad del evento. El potencial de usar fragmentos de cotiledón procedentes de embriones somáticos primarios y embriones somáticos completos cultivados en un sistema de inmersión automático en medio de cultivo líquido, es un avance para la multiplicación de genotipos con caracteres deseables.
AGRADECIMIENTOS
Al INIFAP por su financiamiento a través del proyecto: Estudios para la multiplicación y conversión de embriones somáticos de cacao a plantas porinmersión temporal, con N°SIGI: 183062116.
LITERATURA CITADA
Alejandro-Lázaro A, Azpeitia-Morales A, Sáenz-Carbonell L, Mirafuentes Hernández F (2015) Embriogénesis somática secundaria en el genotipo de cacao (Theobroma cacao L.) INIFAP 1 y su descripción histológica. Nova Scientia 7: 398-417.
Albarrán J, Bertrand B, Lartud M, Etienne H (2005) Cycle characteristics in a temporary immersion bioreactor affect regeneration, morphology, water and mineral status of coffe (Coffea arabica) somatic embryos.Plant Cell Tissue and Organ Culture 81: 27-36.
Borrone JW, Brown S, Kuhn ND, Motamayor J, Schnell R (2007) Microsatellite markers developed from Theobroma cacao L. expressed sequence tags. Molecular Ecology 7: 236-239.
Buffard-Morel J, Verdeil JL, Pannetier C (1992) Embryogenése somatique du cocotier (Cocos nucifera L.) à partir d’explants foliaires: étude histologique. Canadian Journal of Botany 70: 735-741.
Damiano C, La Starza SR, Monticelli S, Gentile A, Caboni E, Frattarelli A (2005) Propagation of Prunus and Malus by temporary immersion. In: Liquid culture Systems for in vitro plant propagation, AnneKathrine Hvoslef-Eide, Walter Preil. pp: 243-251.
Driver JA, Kuniyuki AH (1984) in vitro propagation of Paradox walnut rootstock. HortScience 19: 507-509.
Eskes B (2000) Introductory notes. International workshop on new technologies and cocoa breeding. Kota Kinabalu, Malaysia. pp: 8-11.
García-Gonzáles R, Quiroz K, Carrasco B, Caligari P (2010) Plant tissue culture: Current status, opportunities and challenges. Ciencia e Investigación Agraria 37:5-30.
Komamine A, Murata N, Nomura K (2005) Mechanisms of somatic embryogenesis in carrot suspensión cultures - morphology, physiology, biochemistry, and molecular biology. in vitro Cellular &. Developmental Biology - Plant 41: 6-10.
Lanaud C, Fouet O, Clément D, Boceara M, Risterucci A, Surujdeo-Maharaj MS, et al. (2009) A meta QTL analysis of disease resistance traits of Theobroma cacao L. Molecular Breeding 24: 361-374.
Li Abdoulaye ZT, Maximova S, Guiltinan M (1998) Somatic embryogenesis and plant regeneration from floral explants of cacao (Theobroma cacao L.) using thidiazuron. in vitro Cellular &. Developmental Biology- Plant 34: 293-299.
Martinelli L, Candioli E, Costa D, Poletti V, Rascio N (2001) Morphogenic competence of Vitis rupestris S. secondary somatic embryos with a long culture history. Plant Cell Reports 20: 279-284.
Mathieu LMA, Blervacq AS, David H, Hawkins S, Neutelings G (2006) Germin-like genes are expressed during somatic embryogenesis and early development of conifers. Plant Molecular Biology 61: 615-627.
Maximova SN, Florez S, Shen X, Niemenak N, Zhang Y, Curtis W, et al. (2014) Genome-wide analysis reveáis divergent patterns of gene expression during zygotic and somatic embryo maturation of Theobromacacao L., the chocolate tree. BMC Plant Biology 14: 1-17.
Maximova SN, Ann Young, Pishak S, Guiltinan MJ (2008) Field performance of Theobroma cacao L. plants propagated via somatic embryogenesis in vitro Cellular &. Developmental Biology - Plant 44: 487- 493.
Minyaka E, Niemenak N, Sangare A, Omokolo FND (2008) Characterization of leafy cotyledonl-like during embryogenesis in Theobroma cacao L. Planta 227: 853-866.
Pérez-Nuñez MT, Chan JL, Sáenz L, Gonzáles T, Verdeil JL, Oropeza C (2006) Improved somatic embryo-genesis from Cocos nucífera (L) plumule explants. in vitro Cellular &. Developmental Biology - Plant 42: 37-43.
Salvador N, Espinoza E, Rojas JC (2005) Manual del cultivo de cacao blanco de Piura. Proyecto Norte emprendedor. Lima, Perú. 60p.
Tan C L, Furtek DB (2003) Development of an in vitro regeneration system for Theobroma cacao from mature tissues. Plant Science 164: 407-412.
Urrea T ruj i lio Al, Atehortúa Garcés L, Gallego Rúa AM (2011) Regeneración vía embriogénesis somática de una variedad colombiana élite de Theobroma cacao L. Revista Colombiana de Biotecnología XIII: 39-50.
Watt MP (2012) The status of temporary immersion system (TIS) technology for plant micropropagation. African Journal of Biotechnology 11: 14025-14035.
Yang X, Zhang X (2010) Regulation of somatic embryogenesis in higher plants. Critical Reviews in Plant Science 29: 36-57.
Zahid RM, Das S, Prasad SM, Shankar SP (2013) Somatic embryogenesis in Monocots. In: Somatic embryogenesis and gene expression. Aslam J, Srivastava PS, Sharma MP (Ed.) Narosa. pp: 18-34.
Ziv M (2005) Simple bioreactors for mass propagation of plants, In: Hvoslef-Eide AK, Preil W (Eds). Liquid culture Systems for in vitro plant propagation. Springer, Berlin. pp: 79-94.