Artículo científico
Diversidad de hongos silvestres comestibles del cerro El Pinal, municipio de Acajete, Puebla, México
Diversity of edible wild mushrooms of the hill El Pinal, municipality of Acajete, Puebla, Mexico
1Ricardo I Pérez-López, 3Gerardo Mata, 2Agustín Aragón García, 2Daniel Jiménez García, 2*Omar Romero-Arenas
1 Posgrado en Manejo Sostenible de Agroecosistemas, ICUAP-Benemérita Universidad Autónoma de Puebla (BUAP).
2 Centro de Agroecología, Instituto de Ciencias, ICUAP- BUAP.
*biol.ora@hotmail.com
3 Instituto de Ecología, A. C., Carretera antigua a Coatepec, No. 351, CP 91070, Xalapa, México.
Artículo recibido el 26 de septiembre de 2014
Aceptado el 20 de febrero de 2015
RESUMEN
El estado de Puebla es rico en biodiversidad fúngica, sin embargo hacen falta estudios para determinar la magnitud de la misma. El presente trabajo tuvo como objetivo realizar un estudio de riqueza de hongos comestibles del cerro El Pinal que se ubica en el municipio de Acajete, Puebla. Se recolectaron 86 especímenes de macromicetos de junio a octubre del 2013. Se identificaron 25 especies pertenecientes a 2 clases, 8 órdenes, 17 familias y 19 géneros. De las 25 especies encontradas, 19 pertenecen a los Basidiomycetes y 6 a los Ascomycetes. La proporción por familias muestra en primer lugar a Ramariaceae y Tricholomataceae, ambas con 16 %, seguido de Amanitaceae y Cantharellaceae con 11 %; estas cuatro familias abarcan 54 % del total de especies encontradas para la clase Basidiomycetes, mientras que para la Ascomycetes, la familia mejor representada fue Helvellaceae con el 50 % del total de las especies. La riqueza de especies fue analizada bajo 5 estimadores: ICE, Chao 2, Jack 1, Jack 2 y Bootstrap, los cuales indican una completitud del inventario mayor al 90 %. La curva de acumulación de especies llegó a una asíntota, demostrando que el muestreo fue suficiente para determinar la biodiversidad de hongos silvestres comestibles del cerro El Pinal. Todas las especies identificadas son de alto valor gastronómico, por lo que se requieren estudios sobre su utilización y las prácticas de manejo de este recurso forestal no maderable.
Palabras clave: Aprovechamiento, Bosque de Encino, Macromicetos, Riqueza de especies, Taxonomía
ABSTRACT
The Puebla state is rich in fungal biodiversity, however studies are needed to determine its magnitude of. This research aimed to conduct a richness study of edible wild mushrooms of the hill "El Pinal" in the municipality of Acajete, Puebla. There were collected 86 specimens of macrofungi during the months of June to October in 2013, it identified 25 species belonging to 2 classes, 8 orders, 17 families and 19 genera. Of the 25 species found, 19 belong to Basidiomycetes and 6 belong to Ascomycetes. The proportion of families shows first to Ramariaceae and Tricholo-mataceae, both with 16 %, followed by Amanitaceae and Cantharellaceae with 11 %; these four families contain 54 % of all species found to Basidiomycetes, while for Ascomycetes, best represented family was Helvellaceae with 50 % of all species. The Species richness was analyzed under five estimators: ICE, Chao2, Jackl, Jack2 and Bootstrap, which indicate a greater inventory of 90 %. The species accumulation curve reached an asymptote, demonstrating that sampling was sufficient to determine the biodiversity of edible wild mushrooms of the hill "El Pinal". All species identified in this research are gastronomic value but this information can only be complemented by studies on the use and management practices of this non-timber forest resource.
Key words: Achievement, Oak Forest, Macromycetes, Species Richness, Taxonomy
INTRODUCCIÓN
Los hongos son considerados como el segundo grupo más grande de organismos en la biosfera, después de los artrópodos. Hawksworth (2001) menciona que existe una relación de 6:1, basada en la estimación de plantas vasculares y de hongos, dando como resultado alrededor de 1 500 000 especies en el mundo, de las cuales menos del 5 % ha sido descrito y reportado (Hawksworth 1991, 1993, Hawksworth y Mueller 2005). Estudios recientes basados en habitas potenciales para el desarrollo de especies fúngicas con métodos moleculares, predisponen una alta tasa de acumulación de nuevas especies, con estimación de 3 a 5 millones de especies (Hawksworth y Rossman 1997, O’Brien et al. 2005, Blackwell 2011). Actualmente han sido descritas 300 000 taxa, con cerca de 10 000 especies de macromicetos, de ésta cantidad, alrededor de 2 000 se consideran comestibles (Rossman 1994).
Dadas las características biogeográficas que posee México, se estima que hay alrededor de 200 000 especies de hongos, de las cuales solamente se conoce alrededor del 4 % (Guzmán 1998a, 1998b). Los estudios realizados en México en materia de macromicetos son muy escasos en comparación con otros países, aun cuando la riqueza fúngica en dichos países es menor que en la región tropical de México (Hawksworth 1991, Guzmán et al. 1997).
El manejo, conservación y producción de los hongos, ofrece beneficios agronómicos, alimentarios y medicinales (Guzmán 1994). La distribución geográfica de los hongos comestibles comprende todo el ámbito nacional, pero sólo se dispone de información de 28 estados, siendo los estados con mayor diversidad Puebla, Estado de México, Veracruz, Michoacán y Oaxaca (Villarreal 1995); mientras que los estados en donde se han realizado más estudios son el Estado de México, Veracruz, Hidalgo, Michoacán y Morelos (Villarreal y Pérez-Moreno 1989). El número exacto de especies nativas de hongos comestibles en el estado de Puebla se desconoce (Pellicer-González et al. 2002, Pérez-Moreno et al. 2008). Sin embargo, Puebla tiene características propicias para figurar dentro de los de mayor diversidad, ya que sus bosques cuentan con 770 000 ha, de las cuales 190 000 tienen potencial para el aprovechamiento forestal maderable (Romero 2014). En cuanto a producción forestal, la entidad tiene una superficie total bajo manejo de 84 000 ha, correspondiendo 93.2 % a coniferas, 6.4 % a latifoliadas y 0.4 % a otras especies (Mallén et al. 2005). El desarrollo de estudios sobre la riqueza, taxonomía y ecología de especies de macromicetos con potencial gastronómico, presenta una alternativa para la implementación de diversas acciones encaminadas a la conservación y aprovechamiento sustentable de la biodiversidad. Por lo anterior, el objetivo general de la presente investigación fue conocer la riqueza y distribución fúngica del cerro El Piñal del municipio de Acajete, en el estado de Puebla.
MATERIALES Y MÉTODOS
Área de estudio y recolección de los ejemplares
La recolección de macromicetos silvestres comestibles se realizó en el cerro El Piñal, ubicado en el municipio de Acajete, Puebla, que se localiza entre los 19° 00' y 19° 12' LN, y los 97° 50’ y 98° 01' de LO; a una altitud de entre 2 280 y 3 200 msnm. Colinda al norte con el municipio de Tepatlaxco de Hidalgo, parte del estado de Tlaxcala y el municipio de Nopalucan; al este con los municipios de Nopalu-can y Tepeaca; al sur con los municipios de Tepeaca, Cuautinchán y Amozoc; al oeste con los municipios de Amozoc y Tepatlaxco de Hidalgo (Figura 1). Presenta un clima templado subhúmedo con lluvias en verano C(w2), y temperatura media entre 6 y 16 °C, precipitación pluvial media anual de 772.3 mm y área boscosa de 2 190 ha (INEGI 2009).
Las recolectas de los ejemplares se realizaron en el periodo de lluvias de la región, que comprendió de junio a octubre del 2013. Se realizaron 2 muéstreos mensuales, con 10 transectos de 3 x 100 m cada uno, tomando en cuenta las veredas por donde se trasladan los pobladores (Figura 2), las recoletas fueron realizadas a lo largo de los transectos a un máximo de 3 m de distancia a los lados del camino principal.
Procesamiento e identificación de los ejemplares
Para cada espécimen recolectado se tomaron los siguientes datos: nombre del colector, fecha, localidad, sustrato, coordenadas UTM; en el píleo: tamaño, forma, margen, color, textura, ornamentación; en las láminas: unión, frecuencia, borde; en el estípite: forma, color, consistencia y características de la carne, se tomaron fotografías de cada ejemplar y se colocaron en bolsas de plástico herméticas con sus respectivas etiquetas (Largent 1973, Cifuentes et al. 1986, Largent y Thiers 1986, Delgado et al. 2005). La recolección de basidiomas de los ascomicetes, se realizó en ejemplares completos, evitando aquellos en proceso de pudrición o decolorados por las condiciones climáticas, se utilizaron las guías de colores de Munsell (1975) y, Kornerup y Wanscher (1978). Las muestras se trasladaron al Laboratorio de Fitopatología y Biotecnología de Hongos del Centro de Agroecología del Instituto de Ciencias de la Benemérita Universidad Autónoma de Puebla (BUAP) para su posterior análisis. Para todas las muestras se realizaron preparaciones temporales con KOH a 5 % y reactivo de Melzer, de cortes transversales y longitudinales de las diferentes partes del esporoma, a los que se les evaluaron las siguientes características microscópicas: tamaño, forma y color de las esporas, basidios, cistidios, setas e hifas; también se observó el tipo de sistema hifal y la presencia de fíbulas (Largent et al. 1986). Se midieron entre 25 y 30 esporas, basidios y cistidios, a los que se les determinó el largo y el ancho de cada estructura, con el programa Piximétre 5.3. Para la identificación taxonómica de los ejemplares se utilizaron claves de identificación (Largent 1973, Guzmán 1979, 1989, Largent y Watling 1986, Largent y Baroni 1988), y las claves taxonómicas Mushroom Expert (Kuo 2007).
Para el arreglo taxonómico de los ejemplares así como su correcta revisión nomenclatural, se siguió la clasificación propuesta por el Index Fun-gorum (CABI 2012) y el Catálogo de Autoridades Taxonómicas de los Hongos de México (CONABIO 2008). Una vez concluida la identificación de los hongos, estos se colocaron deshidratados en bolsas de plástico selladas donde se les agregó su información asociada, pasando a formar parte de la Colección de Hongos silvestres (HOUAP) del Centro de Agroecología del Instituto de Ciencias de la BUAP.
Parámetros para la evaluación de la diversidad
Para estimar la riqueza observada y la com-pletitud del inventario de especies de hongos silvestres comestibles del cerro El Piñal se ejecutó un análisis combinado de estimadores no paramétricos para esfuerzos de muestreo crecientes, debido a que es la forma más directa de inferir la biodiversidad de un sitio (Dale y Beyeler 2001, Sarkar 2002, Magurran 2004, González-Oreja et al. 2010b). Se calcularon curvas de acumulación de especies estimadas según los estimadores no paramétricos (Chazdon et al. 1998, Colwell 2006) de cobertura, basado en la incidencia (ICE), estimador Chao 2, estimadores de Jackknife de primer y segundo orden (Jack 1 y Jack 2) y estimador Bootsrap. Se eliminó el posible efecto del orden en el que se añaden las muestras a la curva mediante remuestreo aleatorio, en todos los cálculos se usaron 100 aleatorizaciones con reemplazamiento (Colwell y Coddington 1994). Todas las estimaciones se realizaron con el programa EstimateS vers. 9.1.0 (Colwell 2006); los resultados de todas las reordenaciones aleatorias se exportaron a un archivo de Microsoft Office Excel para evaluar los resultados de cada estimador. La estructura del modelo de acumulación de especies asintótico utilizado fue el de Chapman-Richards:
S(x) = a(1-exp(-bx))c
Dónde S(x)= Riqueza observada acumulada (Sobs; valor medio obtenido tras 100 remuestreos con reemplazamiento), mientras que la variable independiente (x) fue el esfuerzo de muestreo, y a, b, c y el número de parámetros del modelo.
RESULTADOS
Relación y descripción de las especies fúngicas del cerro El Piñal
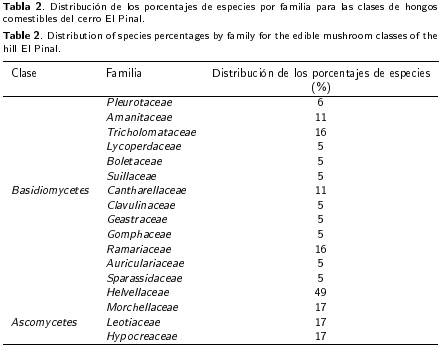
Se recolectaron 86 especímenes de basidio-carpos de hongos silvestres, de los cuales 25 especies se identificaron como hongos comestibles, pertenecientes a 2 clases, 8 órdenes, 17 familias y 19 géneros. De las 25 especies identificadas, 19 pertenecen a los Basidiomycetes y 6 a los Ascomycetes. Las familias presentes de la clase Basidiomycetes fueron: Pleuroteaceae, Amanitaceae, Tricholomataceae, Lycoperdaceae, Boletaceae, Suillaceae, Cantharellaceae, Clavulinaceae, Geastraceae, Gomphaceae, Ramariaceae, Auriculariaceae y Sparassidaceae; mientras que las familias pertenecientes a la clase Ascomycetes fueron: Helvellaceae, Pezizaceae, Morchellaceae, Leotiaceae e Hypocreaceae (Tabla 1).
Pleurotus pulmonarius (Fr) Quél., 1872. Píleo: de 6 cm, convexo a ligeramente plano, húmedo en fresco, coloración blanquecina a café claro. Láminas cerradas, decurrentes blanquecinas. Estípite: ausente. Esporas: de 8 - 10 x 3.5 - 5 µ, cilindricas, depósito blanco. Hábitat: gregario sobre madera en descomposición.
Amanita caesareae (Scop) Pers., 1801. Píleo: de 2.3 - 7.2 cm, de color anaranjado con tonalidades amarillentas, margen estriado, convexo. Láminas lisas, libres, de color amarillo claro. Estípite: de 30 - 65 x 3 - 9 cm, con color a las láminas, fibroso, central, con presencia de un anillo del mismo color de las láminas, presencia de volva blanquecina y membranosa. Esporas de 9 - 12 x 6 - 7 µ, elipsoidales, no amiloides, esporada blanca. Hábitat: solitario en bosque de encino.
Amanita vaginata (Bull) Lam, 1783. Píleo: de 6.3 - 8.2 cm, de color gris a café claro, margen estriado, convexo, ligeramente acampanado. Láminas lisas, libres, blancas. Estípite: de 8 - 12 x 1 - 2 cm, blanco, fibroso, central, presencia de volva blanquecina y membranosa. Esporas de 8 - 10 x 6 - 7 µ, elipsoidales, no amiloides, esporada blanca. Hábitat: solitario.
Gymnopus dryophilus (Bull) Murrill. Homónimo, Colibia drygophila. Píleo: de 2 - 4 cm, ocre-anaranjado, glabro, convexo o ligeramente extendido. Láminas blanquecinas, juntas. Estípite: de 6 - 7.5 x 0.5 cm, blanco y liso. Esporas: de 5.5 - 6 x 2.5 - 3 µ, elípticas, esporada blanca. Hábitat: humícola y gregario.
Hygrophorus chrysodon (Batsch) Fr., 1838. Píleo: de 4 - 6 cm, convexo a ligeramente aplanado, superficie húmeda, blanco con manchas amarillas. Láminas blancas, adheridas, cerosas, ligeramente separadas. Estípite de 5 - 6 x 1 - 1.5 cm, con color al píleo, presencia de manchas amarillentas. Esporas: de 7 - 10 x 3.5 - 5 µ, lisas, elípticas, no amiloides. Hábitat: algo disperso en suelos con abundante materia orgánica.
Hygrophorus russula (Schaeff) Kauffman, 1918. Píleo de 5 - 8 cm, convexo a plano, ligeramente glutinoso y f¡briloso, margen enrollado y liso, color rojizo a rosado. Estípite de 3 - 6 x 1 - 3.5 cm, más o menos con la misma coloración del sombrero, ligeramente fibriloso. Esporas: de 6.2 - 7.4 x 3.5 -4.5 µ, elípticas, lisas, esporada blanca. Sustrato y hábitat: terrícola y disperso.
Lycoperdon perlatum Pers., 1796. Con forma globosa, presenta un área prominente que asemeja a un estípite, el diámetro de la parte más ancha es de 5 cm y con 4 - 7 cm de altura, está cubierto con unas pequeñas prominencias semejantes a espinas pulverulentas que son fácilmente desprendibles, color blanquecino y marrón, maduro presenta una abertura apical. Esporas de 3.5 - 4.5 µp redondas y ornamentadas. Sustrato y hábitat: humícola y gregario.
Boletus edulis Bull., 1782. Píleo: de 8 -12 cm, convexo a ligeramente convexo, húmedo al tacto, liso, color marrón y ligeramente anaranjado. Superficie de los poros: blanquecina a amarillenta, cuando está maduro contiene de 2 a 3 poros por milímetro. Tubos: de 3 cm de profundidad. Estípite: de 10 - 15 cm de largo y 4 cm de grosor, ligeramente reticulado, sobretodo en la parte superior, color blanquecino a marrón en algunas zonas. Esporas: de 13 - 18.5 x 4 - 6.5 µ, esporada marrón oliváceo. Hábitat: solitario y humícola.
Sillus brevipes (Peck) Kuntze, 1898. Píleo: de 5 - 8 cm, convexo, glutinoso, color marrón rojizo, margen ligeramente incurvado. Superficie de poros: amarillo pálido, oliváceo, de 1 a 2 poros circulares por milímetro. Tubos: de 1 cm de profundidad. Estípite: 2 - 5 x 1 - 3 cm, con color a la superficie de los poros. Esporas: de 7 - 9.5 x 2 - 3 µ, lisas y subfusiformes, esporada marrón canela. Pleurocistidios y queilocistidios de 50 x 10 µ ligeramente marrones. Hábitat: solitario o en grupos, en asociación con pinos.
Cantharellus cibarius Fr., 1821. Píleo: de 1 - 6 cm, más o menos convexo volviéndose plano o ligeramente involuto, margen irregular y recurvado, liso a ligeramente fibriloso, color amarillo pálido. Láminas: presencia de láminas falsas con apariencia de inervaciones decurrentes, con color al píleo aunque ligeramente más pálidas. Estípite: de 2.4 -6x1-2 cm, de forma variable, y de coloración como la parte himenial. Esporas: de 7 - 10.5 x 4 - 5.5 µ, lisas, elípticas, no amiloides. Esporada: amarillo pálido. Hábitat: gregario, en suelos con abundante hojarasca.
Craterellus cornucopioides (L) Pers., 1825. Cuerpo fructífero: de 2 - 7 cm de ancho por 10 cm de altura, tubular, con margen enrollado cuando es joven, de carne delgada, la parte interna es de una coloración obscura con tonos grises y con la presencia de escamas pálidas en su superficie, la parte externa es lisa y con ligeras enervaciones cercanas a la parte del margen, de color cenizo. Esporas: de 8.4 - 13.2 x 5.5 - 5.9 µp, lisas y elípticas. Hábitat: gregario, casi enterrado sobre la hojarasca.
Clavulina coralloides (L) J. Schröt., 1888. Cuerpo fructífero: de 2 - 10 x 3 - 5 cm, ramificaciones juntas, de 0.2 - 0.5 cm de anchura, de color blanquecino a grisáceo, la base es delgada de aproximadamente 0.5 cm de anchura y hasta 3 cm de largo. Esporas: de 7 - 11 x 6.5 - 10 µ, basidios clavariformes con 2 esterigmas. Hábitat: solitario a gregario, sobre suelo.
Geastrum triplex Jungh., 1840. Cuerpo fructífero: en forma de pelota con una abertura prominente, de 2 - 4 cm, se desgaja en cuatro o cinco pétalos de coloración café claro, se ennegrece con la edad, la parte interna de los gajos suele desgarrarse formando lo que pareciera una tercera capa. Esporas: de 4 - 4.5 µ, globosas, con una ornamentación de espinas, coloración marrón. Hábitat: disperso al pie de encinos.
Clavariadelphus truncatus (Quél) Donk, 1933. Cuerpo fructífero: de 3 - 15 cm de altura y cerca de 3 cm de ancho en la parte apical, clavariforme, al principio es liso y con la edad se vuelve rugoso en su superficie, rosado anaranjado, cuando maduro se obscurece y torna marrón, de carne blanquecina. Esporas: de 8.3 - 11.6 x 4 - 5.5 µ, lisas. Hábitat: solitario a gregario, abundante y en asociación clara con encino.
Ramaria flava (Schaeff) Quél., 1888. Cuerpo fructífero: de 4 - 10 x 5 - 10 cm, base pobremente definida, ramificaciones juntas, ramificaciones elongadas y orientadas verticalmente, de coloración pálida, blanquecina y en ocasiones ligeramente rosada, se oscurece con el maltrato, la base es muy poco diferenciada, blanquecina, con presencia de ri-zomorfos. Esporas: de 7.5 - 10.5 x 3.5 - 4.5 µ, elípticas, rugosas, presencia de hifas de pared gruesa. Hábitat: solitario a gregario, sobre suelos ricos en materia orgánica.
Ramaria stricta (Pers) Quél., 1888. Cuerpo fructífero: de 4 - 10 cm alto x 3 - 8 ancho cm, forma coraloide muy ramificada, tronco delgado de color amarillento formado por numerosas ramas verticales, estrechas, largas y paralelas y extremos bifurcados en puntas cortas. Himenio liso y mate, que cubre toda la superficie ramosa. Carne entre blanca y pardo rosado. Olor débil y agradable, pero sabor amargo. Esporas: de7 - 9 x 4 - 5 µ, anchamente elípticas, verrugosas, casi lisas, con apícula lateral. Hábitat: frecuentemente encontrada sobre madera muerta, ramas semienterrada, aislada o en grupo.
Ramaria botrytis (Pers) Ricken, 1918. Cuerpo fructífero: de 7 - 15 x 6 - 20 cm, cuerpo macizo, ramificaciones densas, basalmente gruesas, juntas y cortas, de coloración blanquecina en la base y en la parte terminal rosadas a purpura o rojizas, la base es bastante gruesa, blanquecina o ligeramente marrón. Esporas: de 14.5 - 16 x 4 - 5.5 µ, subfusiformes, ornamentadas, basidios de 4 esterigmas. Hábitat: solitario a gregario, sobre suelos ricos en materia orgánica.
Auricularia auricula-judae (Fr) Quél., 1886. Cuerpo fructífero: irregular, típicamente con forma de oreja, de 1 - 4 cm en forma de concha o pétalo, consistencia gelatinosa, coloración de café a marrón obscuro, superficie venosa y suave. Esporas: de 12.5 - 18.4 x 4.3 - 7.6 µ, fusiformes, lisas. Esporada: blanca. Hábitat: solitario e imbricado, sobre madera en descomposición.
Sparassis crispa (Wulfen) Fr., 1821. Cuerpo fructífero: de 10 - 20 cm, ramificaciones en forma de láminas que provienen de una base común, coloración blanquecina. Esporas: de 5.4 - 7 x 4 - 5.5 µ, lisas, ligeramente elípticas, no amiloides. Hábitat: solitario e imbricado sobre la base de encinos principalmente.
Helvella crispa (Scop) Fr., 1822. Sombrero: de 1 - 5 cm en forma de silla de montar, superficie glabra y con presencia de pequeñas venaciones, coloración blanquecina, o color hueso, superficie interna ligeramente fibrilosa, de color un poco más obscuro que la parte exterior. Pie: de 2 - 8 x 1 -2 cm, de color blanquecino más pálido que la parte superior, con presencia de venaciones y huecos a lo largo de todo el pie. Esporas: de 16.5 - 20.2 x 10.4 - 13.7 µ, elípticas, lisas, con presencia de gotas internas, ascosporas de 8 esporas, paráfisis clavadas, hialinas de 5.2 - 12.2 µ de ancho. Hábitat: disperso y terrícola bajo árboles de encino.
Helvella lacunosa Afzel., 1783. Sombrero: de 5 - 8 cm en forma de silla de montar, superficie glabra y con presencia de pequeñas venaciones, coloración marrón obscuro a negro, superficie interna lisa, de color gris o tonos marrones grisáceos. Pie: de 2 - 10 x 1 - 3 cm, de color blanquecino cuando joven, se torna obscuro y puede llegar a ser grisáceo o negro, con presencia de venaciones y huecos a lo largo de todo el pie. Esporas: de 14.5 -21.2 x 10.4 - 13.7 µ, elipsoides, lisas, con presencia de gotas internas, ascosporas de 8 esporas, paráfisis clavadas, hialinas de 2.5 - 10.2 µ de ancho. Hábitat: disperso y terrícola regularmente asociado con coniferas.
Helvella cupuliformis Dissing &. Nannf., 1966. Sombrero: de 1 - 4 cm en forma de copa, superficie lisa y coloración gris y en ocasiones marrón, superficie interna lisa, de color gris o tonos marrones grisáceos, se torna más pálida hacia el pie. Pie: de 1 - 3 x 0.5 - 0.8 cm, de color blanquecino a café. Esporas: de 16.3 - 20.2 x 11.4 - 12.7 µ, elipsoides, lisas, con presencia de gotas internas, paráfisis con contenido granular de color café, hialinas a café, de 4 - 8 µ de ancho. Hábitat: disperso a gregario, terrícola, en suelo de bosque de encino.
Morchella esculenta (L) Pers., 1794. Sombrero: de 5 - 8 cm en forma de colmena, con numerosos alveolos, de coloración marrón u ocre. Pie: de 2 - 4 x 1.5 - 2 cm, de color blanquecino. Esporas: de 16.3 - 20.2 x 11.4 - 13.7 µ, elipsoides, lisas, con presencia de bordes granulosos. Hábitat: disperso a gregario, terrícola, en suelo de bosque de encino, comestible.
Leotia lubrica (Scop) Pers., 1797. Sombrero: de 1 - 4 cm, de forma variable, convoluto, liso, viscoso al tacto, margen enrollado, coloración café amarillento, a oliváceo. Pie: de 2.5 - 6 x 0.8 - 1 cm, liso, viscoso, con color al sombrero aunque más pálido. Esporas: de 16.6 - 24.5 x 4 - 5.5 µ, subfusiformes, septadas, paráfisis filiformes de 1 - 2 µ de ancho. Hábitat: cespitoso, terrícola, en suelo de bosque de encino.
Hypomyces lactifluorum (Schwein) Tul. &. C. Tul., 1860. Cuerpo fructífero: parasito que recubre en su totalidad los basidiomas de especies pertenecientes al género Russula y Lactarius, los torna completamente a una coloración rojizo-anaranjado. Esporas: de 35 - 50 x 4 - 5.5 µ, alargadas, septadas. Hábitat: disperso, fungícola, comestible.
La proporción de familias dentro de la clase Basidiomycetes (Tabla 2), muestra en primer lugar a la familia Ramariaceae y Tricholomataceae, ambas con 16 % seguido de Amanitaceae y Cantharellaceae con el 11 %, tan solo estas cuatro familias abarcan 54 % del total de especies encontradas para la clase Basidiomycetes; mientras que para la clase Ascomycetes, la familia mejor representada fue Helvellaceae con 50 % del total de las especies, todas las demás familias ocupan un porcentaje inferior.
El bosque de encino presentó un punto máximo de incidencia de especies de hongos comestibles en el mes de septiembre. Las especies que presentaron mayor recurrencia durante los meses de colecta fueron: Auricularia auricula-judae que se encontró en todos los meses de recolecta, mientras que Clavariadelphus truncatus, Lycoperdon perlatum y Ramaria flava se encontraron en cuatro de los cinco meses muestreados.
Análisis de la riqueza de especies de hongos comestibles
La riqueza estimada de macromicetos con los índices de ICEICE, Chao 2, Jack 1, Jack 2 y Bootstrap (Figura 3), mostraron un porcentaje de completitud del inventario superior al 90 %. Los estimadores sugieren, en la mayoría de los casos una alta completitud de muestreo de hongos silvestres comestibles a partir del quinto muestreo. La curva acumulativa de especies alcanza una asíntota en el valor Sobs = 33. El resultado del estimador más bajo respecto a lo observado fue Jack 2 con 31 especies, y el más alto fue Bootstrap con 34 especies,la asíntota para el resto de los estimadores coincidió en 33 especies de hongos comestibles para el cerro El Piñal.
DISCUSIÓN
Identificación y presencia de los ejemplares
Todas las especies identificadas en el presente estudio han sido reportadas como comestibles (Villarreal y Pérez Moreno 1989, Montoya et al. 2001, 2004, FAO 2005). La clase Basidiomycetes mostró 4 familias importantes: Ramariaceae, Tricholomataceae, Amanitaceae y Cantharellaceae, en las cuales se integran algunos de los taxones más frecuentemente citados para México (Herrera y Guzmán 1961, Montoya et al. 2004). La familia Ramariaceae estuvo representada con tres especies del género Ramaria (R. flava, R. stricta y R. botrytis), donde R. flava tuvo una incidencia en cuatro de los cinco meses de recolecta además de que su abundancia fue evidente. La familia Tricholomataceae, donde se identificaron 3 especies, una del género Gymnopus: G. dryophilus y dos del género Hygrophorus: H. chrysodon e H. Russula. Los tonos vi naceos para H. russula y los amarillentos para H. chrysodon las hacen inconfundibles respecto a otras especies. Para la familia Amanitaceae es importante mencionar que sus dos especies identificadas tiene alto valor culinario y pertenecen al género Amanita. Por lo que se refiere a A. caesarea y A. vaginata se encontraron en abundancia en los meses de agosto y septiembre, resultados que son similares a los reportados por Romero-Arenas et al. (2009) para la sierra norte del estado de Puebla. En la familia Cantharellaceae se identificaron dos especies, Cantharellus cibarius y Craterellus cornucopioides, ambos tienen formas similares pero de coloración muy distinta, la primera es conspicua, ya que se colecto en varias salidas en abundancia. Aunque las familias Auriculariaceae, Lycoperdaceae y Gomphaceae dentro de la clase Basidiomycetes presentaron las especies Auricularia auricula-judae, Lycoperdon perlatum y Clavariadelphus truncatus, las cuales se encontraron en cuatro de los cinco meses de muéstreos. Dentro de la clase Ascomycetes la familia Helvellaceae fue la mejor representada con tres especies comestibles del género Helvella (H. crispa, H. cupuliformis y H. lacunosa), sin embargo, para la especie Morchella esculenta sólo se colectaron en el mes de septiembre, cabe destacar que se recolectó a 2 800 msnm, en los límites del bosque de encino con la vegetación de coniferas. Esta especie es muy apreciada en la región (Montoya et al. 2001).
Se recolectaron 33 especies de macromicetos, de las cuales solo 25 se identificaron, las ocho restantes fueron morfoespecies que pertenecen a los géneros: Lactarius, Boletus y Ramaria. Los muéstreos se realizaron en la vegetación predominante de pino-encino del cerro El Piñal, mientras que en la parte más alta y menos accesible, el tipo de vegetación es de bosque de coniferas, en donde se pueden encontrar hongos comestibles distintos a los encontrados en la vegetación de pino-encino.
Análisis de la riqueza de hongos silvestres comestibles
La función de la acumulación de especies de Chapman-Richards tuvo un coeficiente de determinación (R2) mayor al 99 %, más no se descarta la utilización de otro modelo de función sobre la acumulación de especies. El análisis de riqueza para este estudio indicó un esfuerzo de muestreo suficiente y a partir del quinto muestreo el número promedio de especies observadas se reduce considerablemente disminuyendo la probabilidad de encontrar nuevas especies. La asíntota se alcanza alrededor de S(x) = 33, esto es consistente con los modelos de riqueza de especies, además los estimadores tienen las características descritas por Toti et al. (2000). Al respecto González-Oreja et al. (2010a) mencionan que cuando un inventario está prácticamente completo, las diferencias en las asíntotas estimadas que ofrecen los distintos modelos suelen ser pequeñas como es el caso del presente estudio. Los modelos aquí evaluados, podrían ser utilizados para determinar la riqueza de hongos silvestres comestibles del cerro El Pinal. Sin embargo, aunque se haya completado el inventario de acuerdo con la estimación, eso no indica que sean todas las especies de hongos comestibles en la región, debido a que el inicio de la temporada de recolectas comenzó en el mes de junio. Al respecto Mariaca-Méndez et al. (2001) reportan especies de Agaricus sp., para el Valle de Toluca, México en los meses de abril y mayo, periodo no contemplado es este estudio.
CONCLUSIONES
Se actualizó el conocimiento sobre los hongos comestibles del cerro El Piñal, del municipio de Acajete en el estado de Puebla; se identificaron 25 especies, que pertenecen a 2 clases, 8 órdenes, 17 familias y 19 géneros. El bosque de pino-encino tuvo gran diversidad de hongos comestibles, principalmente en los meses de agosto y septiembre; el análisis de la riqueza de especies fue concluyente, presentando una completitud del inventario mayor al 90 %. Aunque las especies identificadas en la presente investigación han sido reportadas como comestibles, hace falta más investigación etnomicológica en la región, lo que podría arrojar información complementaria sobre la utilización y prácticas de manejo de este recurso forestal no maderable.
LITERATURA CITADA
Blackwell M (2011) The Fungi: 1, 2, 3 ... 5.1 Million Species? American Journal of Botany 98: 426-438.
CABI (2012) Bioscience, CBS &. Landcare Research, https://www.indexfungorum.org. Fecha de consulta 26 de octubre de 2013.
Chazdon RL, Colwell RK, Denslow JS, Guariguata MR (1998) Statistical methods for estimating species richness of woody regeneration in primary and secundary rain forests of Northeastern Costa Rica. In: Dallmeier F, Comiskey JA (eds.) Biodiversity Research, Monitoring and Modeling: Conceptual Background and Old World Case Studies. The Parthenon Publishing Group, Paris. pp: 285-309.
Cifuentes J, Villegas M, Pérez-Ramírez L (1986) Hongos. In: Lot A, Chiang F (comp.) Manual de Herbario. Consejo Nacional de la Flora de México, A.C. México. 142p.
Colwell RK (2006) Estimates: Statistical Estimation of Species Richness and Shared Species from Samples. Version 8. User’s Guide and application, http://purl.oclc.org/estimates. Fecha de consulta 15 de noviembre de 2013.
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society (Series B) 345: 101-118.
CONABIO (2008) Catálogo de autoridades taxonómicas de los hongos (Fungi) de México. Base de datos SNIB-CONABIO. México, http://www.biodiversidad.gob.mx/especies/gran_familia/hongos/docs/Hongos.pdf. Fecha de consulta 24 noviembre 2013.
Dale VH, Beyeler SC (2001) Challenges in the development and use of ecological indicators. Ecological Indicators 1: 3-10.
Delgado A, Villegas M, Cifuentes J (2005) Glosario ilustrado de los caracteres macroscópicos en Basidiomycetes con himenio laminar. Facultad de Ciencias. UNAM, México. 84p.
FAO (2005) Los hongos silvestres comestibles: Perspectiva global de su uso e importancia para la población. Roma, Italia. 156 p. https://www.fao.Org/3/a-y5489s.pdf. Fecha de consulta 28 de septiembre de 2013.
González-Oreja JA, De la Fuente-Díaz AA, Hernández-Santín L, Buzo-Franco D, Bonache-Regidor C (2010a). Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México. Animal Biodiversity and Conservation 33: 31-45.
González-Oreja JA, Garbisu C, Mendarte S, Ibarra A, Albizu I (2010b) Assessing the performance of nonparametric estimators of species richness in meadows. Biodiversity and Conservation 19: 1417-1436.
Guzmán G (1979) Identificación de los hongos comestibles, venenosos, alucinantes y destructores de la madera. Editorial Limusa. México. 451p.
Guzmán G (1989) Hongos. Editorial Limusa. 2a Edición. México. 124p.
Guzmán G (1994) Los hongos y liqúenes en la medicina tradicional Mexicana. En: Argueta A, Cano LM, Rodarte ME (eds.) Atlas de las Plantas de la medicina Tradicional Mexicana, vol. III. Instituto Nacional Indigenista. México. pp: 1427-1487.
Guzmán G (1998a) Inventorying the fungi of Mexico. Biodiversity and Conservation 7: 369-384.
Guzmán G (1998b) Análisis cualitativo y cuantitativo de la diversidad de los hongos en México (Ensayo sobre el inventario fúngico del país). En: HaIffter G (eds.) La diversidad biológica de Iberoamérica II. Acta Zoológica Mexicana. CYTED e Instituto de Ecología de Xalapa. pp: 111-175.
Guzmán G, Bandala VM, Montoya L (1997) An overview on the tropical fungi from Mexico. In: Janardhanan KK, Rajendran C, Natarajan N, Hawksworth DL (eds). Tropical Mycology. Science Publ. Enfield New Hampshire, E.U.A. pp: 115-147.
Hawksworth DL, Rossman A Y (1997) Where are all the undescribed fungi?. Phytopathology 87: 888- 891.
Hawksworth DL (1991) The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycological Research 95: 641-655.
Hawksworth DL (1993) The tropical fungal biota: census, pertinence, prophylaxis, and prognosis. In: Issac S, Frankland JC, Watling R, Whalley AJS (eds). Aspects of Tropical Mycology. Cambridge University Press. Cambridge. pp: 265-293.
Hawksworth DL (2001) The magnitude of fungal diversity: the 1.5 million species estimate revised. Mycological Research 105: 1422-1432.
Hawksworth DL, Mueller GM (2005) Fungal communities: their diversity and distribution. In: Digthon J, White JF, Oudemans P (eds.). The fungal community: its organization and role in the ecosystem. CRC Press. Boca Raton, E.U.A. pp: 27-37.
Herrera T, Guzmán G (1961) Taxonomía y ecología de los principales hongos comestibles de diversos lugares de México. Anales del Instituto de Biología. Universidad Nacional Autónoma de México 32: 33-135.
INEGI (2009) Instituto Nacional de Estadística, Geografía e Informática 2005. Prontuario de información geográfica municipal de los Estados Unidos Mexicanos. Acajete, Puebla, http://www3.inegi.org.mx/sistemas/mexicocifras/datos-geograficos/21/21001.pdf. Fecha de consulta 24 de febrero de 2014.
Kornerup A, Wanscher JH (1978) Methuen handbook of colour. Eyre, London. 252p.
Kuo M (2007) Key to major groups of mushrooms, http://www.mushroomexpert.com/major_groups.html Fecha de consulta 21 de octubre de 2013.
Largent D (1973) How to identify mushrooms to genus, I. Macroscopic Features. Mad River Press, Eureka. 166p.
Largent D, Baroni T (1988) How to identify mushrooms to genus, VI. Modern Genera. Mad River Press. Eureka. 277p.
Largent D, Johnson D, Watling R (1986) How to identify mushrooms to genus, III. Microscopic Features. Mad River Press. Eureka. 148p.
Largent D, Thiers HD (1986) How to identify mushrooms to genus, II. Field identification of Genera. Mad River Press. Eureka. 32p.
Largent D, Watling R (1986) How to identify mushrooms to genus, IV. Keys to Families and Genera. Mad River Press. Eureka. 93p.
Magurran AE (2004) Measuring Biological Diversity. Species abundance distributions over time. Ecology Letters 10: 347-354.
Mallén RC, Vidal GC, Tamarit UJC (2005) El manejo de los bosques templados en Puebla. Criterios e indicadores para evaluar la sustentabilidad. CEÑID, COMEF. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. INIFAP. México. 262p.
Mariaca-Méndez R, Silva LC, Castaños CA (2001) Proceso de recolección y comercialización de hongos comestibles silvestres en el Valle de Toluca, México. Ciencia Ergo Sum 8: 30-40.
Montoya A, Estrada-Torres A, Kong A, Juárez-Sánchez L (2001) Commercialization of wild mushrooms during market days of Tlaxcala, Mexico. Micología Aplicada International 13: 31-40.
Montoya A, Kong A, Estrada-Torres A, Cifuentes J, Caballero J (2004) Useful wild fungi of La Malinche National Park, México. Fungal Diversity 17: 115-143.
Munsell C (1975) Munsell soil color chart. United States Department of Agriculture, Kollmorgen, Maryland. 34p.
O’Brien BL, Parrent JL, Jackson JA, Moncalvo JM, Vilgalys R (2005) Fungal community analysis by large-scale sequencing of enviromental samples. Applied and Environmental Microbiology 71: 5544-5550.
Pellicer-González E, Martínez-Carrera D, Sánchez M, Aliphat M, Estrada-Torres A (2002) Rural management and marketing of wild edible mushrooms in Mexico. In: Sánchez JE, Huerta G, Montiel E (eds). Mushroom biology and mushroom products. UAEM. Cuernavaca, México, pp: 433-443.
Pérez-Moreno J, Martinez-Reyes M, Yescas-Pérez A, Delgado-Alvarado A, Xoconostle-Cázares B (2008) Wild mushroom markets in central Mexico and a case study at Ozumba. Economic Botany 62: 1-12
Romero-Arenas O, Huerta-Lara M, Becerril-Herrera M, Bautista-Calles J, Damian-Huato MA, Tapia-Rojas A, Valencia De Ita MA, Bonilla-Vázquez L (2009) Diversity of Wild Mushrooms in the Commonwealth of Benito Juarez, Tetela De Ocampo; Puebla, Mexico. Research Journal of Biological Sciences 4: 179-186.
Romero CS (2014) La biodiversidad en Puebla Estudio de Estado. Usos de la biodiversidad en el estado de Puebla, el patrimonio forestal de Puebla y su problemática. pp: 243-280.
Rossman AY (1994) A strategy for an all taxa of fungal biodiversity. In: Peng Cl, Chou CH (eds). Biodiversity and terrestrial ecosystems. Institute of Botany. Academy Sinica Monograph Series 14: 169-194.
Sarkar S (2002) Defining "biodiversity": assessing biodiversity. The Monist 85: 131-155.
Toti DS, Coyle FA, Miller JA (2000) A structured inventory of appalachian grass bald and heath bald spider assemblages and a test of species richness estimator performance. Journal of Arachnology 28: 329-345.
Villarreal L (1995) Los hongos silvestres, una alternativa para el manejo integral de los bosques. In: Boege E, García Campos H, Gerez-Fernández P (eds.) Alternativas al manejo de laderas en Veracruz. SE-MARNAT y Fundación Friedrich Ebert. México. pp: 197-201.
Villarreal L, Pérez-Moreno J (1989) Los hongos comestibles silvestres de México, un enfoque integral. Micología Neotropical Aplicada 2: 77-1