Nota Cientifica
ACTIVACIÓN in vitro DE MONOCITOS DE BOVINO CON Lactobacillus casei: PRODUCCIÓN DE ÓXIDO NÍTRICO
in vitro Activation of bovine monocytes with Lactobacillus casei: nitric oxide production
Carlos Ramón Bautista Garfias1*, Astrid Rodríguez Lozano1, Jesús Antonio Álvarez Martínez1, Carmen Rojas Martínez1, Julio Vicente Figueroa Millán1, Mariel Díaz López2, Virginia Guadalupe García Rubio2
1 Centro Nacional de Investigación Disciplinaria en Parasitología Veterinaria del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, CP. 62550/AP. 206, Jiutepec, Morelos, México.*Autor de correspondencia: foto.dibujo@gmail.com, bautista.carlos@inifap.gob.mx
2 Centro Universitario UAEM, Amecameca, Universidad Autónoma del Estado de México.
Nota cientifica recibida el 9 de enero de 2015
Aceptado el 25 de septiembre de 2015
RESUMEN
La presente investigación se llevó a cabo con el objetivo de determinar la producción de óxido nítrico (ON) a partir de monocitos de sangre periférica de bovino, expuestos in vitro a L. casei. Se midió la producción deON cada 24 h durante cuatro días, en cultivos de monocitos de sangre periférica de bovinos de 4 a 6 meses de edad(MDE) (n = 4), 8 MDE (n = 3) y 12 MDE (n = 8), expuestos (E) o no (NE) a Lactobacillus casei. En cada unode los periodos de 24 h se observó una producción significativa (p < 0.01) de ON en los monocitos expuestos encomparación con los no expuestos. La diferencia fue más marcada en los monocitos del grupo 4 a 6 MDE. El usode L. casei puede ser importante para establecer medidas de estimulación de la inmunidad innata en bovinos contraenfermedades producidas por diversos patógenos.
Palabras clave: Monocitos, bovino, activación in vitro
ABSTRACT
This research was carried out with the objective of determine the production of nitric oxide (NO) from monocytes of bovine peripheral blood exposed in vitro to L. casei. The NO production was measured every 24hours during of 4 d in monocyte cultures of bovine peripheral blood of 4 to 6 months of age (MOA) (n = 4), 8 MOA (n = 3), and 12 MOA (n = 8), exposed (E) or not exposed (NE) to Lactobacillus casei. On each of the 24 h periods, asignificant production (p < 0.01) of NO was observed on the monocytes exposed in comparison with those not exposed. The difference was more clear on the monocytes of groups 4 and 6 MOA. The use of L. casei can be important inestablishing stimulation measures for the innate immunity in bovines against diseases produced by arious pathogens.
Key words: Monocytes; bovine; in vitro activation.
INTRODUCCIÓN
Los mecanismos inmunitarios de los bovinos se dividen en innatos y adquiridos. Entre los primeros se encuentran células imprescindibles como son los monocitos y los macrófagos (Adams 1979, Auffray et al. 2009). Los monocitos son células que unen las respuestas inmunitarias innata y adquirida y desempeñan un papel fundamental como células dela primera línea de defensa contra agentes patógenos (Hume et al. 2002). Los monocitos se originan en la médula ósea y son liberados en la circulación sanguinea, donde fluyen por varios días antes de entrar a los tejidos para reemplazar las poblaciones de macrófagos y células dendríticas (Auffray et al. 2009). En estudios previos, se ha demostrado que Lactobacillus casei induce respuestas protectoras contra Babesia microti en ratones, cuando se inocula antes o el mismo día de la infección de confrontación (Bautista-Garfias et al. 2005) y contra B. bovis y B. bigemina en bovinos, cuando la bacteria ácido-láctica se administra junto con la vacuna mixta contra babesiosis bovina, desarrollada por el Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP) del gobierno mexicano, bajo condiciones controladas (Bautista et al. 2008) y también bajo condiciones de campo (Bautista-Garfias et al. 2012). Los mecanismos responsables de la inducción de inmunidad innata, generada por el lactobacilo no se conocen, aunque se sugiere que la estimulación de células como los monocitos y macrófagos por L. casei podría ser uno de los mecanismos (Bautista-Garfias et iosal. 2005).
Por otro lado, el óxido nítrico (ON) es un no radical libre sintetizado a partir de L-arginina por mtres diferentes sintasas-ON (ONS) que exhibe múltiples y complejas funciones biológicas y muchos de sus efectos pueden ser atribuidos principalmente a su potente capacidad oxidativa (Pavenelliy Nogueira 2010). Se sabe que el ON puede modular directa e indirectamente la respuesta inmunitaria por medio de diversos mecanismos tales como el me- usdiar efectos microbicidas de radicales libres derivados del ON, principalmente dentro de macrófagos y otros fagocitos (Pavenelli y Nogueira 2010). Se ha indicado que la producción de óxido nítrico por fagocitos, como los monocitos y macrófagos, es un factor importante en la protección del hospedero contra la infección de diferentes microorganismos patógenos (Fang 1997) entre los que se encuentran parásitos como Giardia (Pavanelli et al. 2010), Trypanosoma cruzi (Pavenelli y Nogueira Silva 2010) y Babesia spp. (Johnson et al. 1996, Bautista et al. 2005). En este sentido, también en investigaciones anteriores, se demostró que los monocitos de sangre periférica de bovino modifican su morfología al ser estimulados in vitro por el lactobacilo (Bautista et al. 2014); sin embargo, no se conoce si dicha bacteria activa monocitos para que produzcan ON. En este contexto, la presente investigación se llevoa cabo con el objeto de determinar la producción de ON a partir de monocitos de sangre periférica de bovino, expuestos in vitro a L. casei.
MATERIALES Y MÉTODOS
Animales experimentales
Se utilizaron bovinos Holstein, de diferentes establos de Amecameca, Estado de México. Secolectó sangre periférica de los siguientes tres grupos: 1) 4-6 meses de edad (MDE) (n = 4), 2) 8MDE (n = 3) y 3) 12 MDE (n = 8).
Lactobacilos
Se usó la cepa ATTCC7469 de Lactobacillus casei (Bautista-Garfias et al. 2001) que se cultivó en medio MRS (Man-Rogosa-Sharpe) a 37 °C durante 18 h. Las bacterias se cosecharon y centrifugarona 5000 X g por 10 min para luego lavar tres vecescon solución salina amortiguadora de fosfatos (pH7.2). Después del conteo viable, la cantidad de L. casei se ajusto a una concentracion de 109 unidades formadoras de colonia (ufc)/mL-1.
Aislamiento de monocitos de sangre periférica
De cada uno de los bovinos se tomó una muestra de 10 a 25 mL de sangre periférica porpunción de la vena yugular en tubos Vacutainer conheparina como anticoagulante, previa asepsia contorundas de algodón y benzal al 70 %. Como un primer paso, los leucocitos se aislaron por medio dela centrifugación de la sangre en un gradiente dedensidad (Ficoll-Paque). Después de cuatro lavados por centrifugación a 450 g durante 15 min se determinó la viabilidad celular por medio de tincióncon azul de tripán y un conteo celular en la cámara de Neubauer. Para luego ajustar a 1x106 células con una viabilidad mayor al 95 % por cada caja de Petride seis cm de diámetro. Los monocitos se aislaron con base en su propiedad de adherirse a superficiesde plástico (Birmingham y Jeska 1980, Goddeeris et al. 1986, Adams y Czuprynski 1990). Las cajasse incubaron durante dos horas en estufa de C02 a 37 °C y luego se lavaron con solución de Vega y Martínez (Vega et al, 1985) para descartar células no adherentes y posteriormente se adicionó el medio de cultivo MF12 conteniendo 30 mMol N-Tris (hydroxymethil)-methyl-2-aminoethanesulfonic acid (TES) Y 26 mM NaHCO3.
Cultivo in vitro
Los tratamientos consistieron en la exposición de los monocitos a L. casei (1x1O8 ufc caja-1), obtenido de acuerdo con Bautista et al. (2001) o a solución salina amortiguadora de fosfatos (pH 7.0) (testigo). Cada tratamiento se realizó por triplicado, manteniendo los cultivos in vitro dentro de una incubadora a 37 °C con una atmósfera C02 por 96 horas y realizando cambios de medio cada 24 horas en cada una de las réplicas. Los sobrenadantes obtenidos se congelaron a -20 °C hasta su uso.
Determinación de óxido nítrico (ON)
El ON se determinó en los sobrenadantes de los cultivos obtenidos cada 24 horas por medio de la adición del reactivo de Griess, utilizando un kit comercial (Quantichrom™ Nitric Oxide Assay Kit, BioAssay Systems, CA, USA), siguiendo las instrucciones del fabricante, para luego leer en un lector de ELISA a 595 nm. Los datos se registraron como NO2 (µg M), para luego realizar el análisis estadístico.
Análisis estadístico
Con los datos obtenidos, se llevó a cabo un análisis de varianza de bloques al azar con el paquete de diseños experimentales de la FAUNL, v.2.5 2 (Olivares, 1994). En cada uno de los grupos de diferentes edades, se comparó la producción de ON a las 24, 48, 72 y 96 horas después del tratamiento, entre células tratadas y no tratadas con L. casei. Asimismo, se comparó el promedio total de la producción de ON entre los tres grupos de edad. Cuando se encontraron diferencias se usó la prueba de Tukey y éstas se considerando como significativas cuando los valores de P fueron < 0.01.
RESULTADOS Y DISCUSIÓN
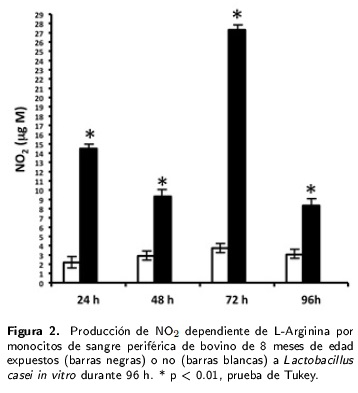
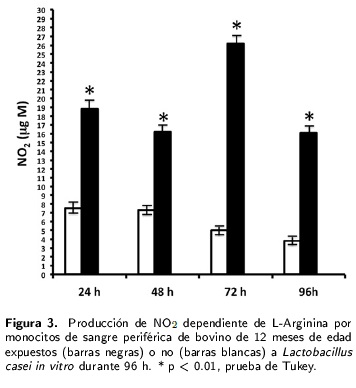
Se observó una diferencia significativa (p < 0.01) en la producción de ON por los monocitos de becerros de 4-6 meses de edad que fue manifiesta desde las 24 h post-exposición a L. casei y que semantuvo a niveles similares a las 48, 72 y 96 h. El promedio mínimo de ON (µg M) fue de 33.2 a las48 h y el máximo de 37.7 a las 96 h post-exposición (Figura 1) en comparación con las células testigo no expuestas al lactobacilo. En el caso de los monocitos de animales de ocho meses de edad se apreció una diferencia significativa (p < 0.01) en la producciónde ON a las 24, 48, 72 y 96 h post-exposición a L. casei. El promedio mínimo de ON (µg M) fuede 8.3 a las 96 h y el máximo fue de 27.3 a las 72 h post-exposición, en comparación con las células testigo no expuestas al lactobacilo (Figura 2). Los monocitos de bovinos de 12 meses de edad también produjeron valores significativamente más altos de ON (µg M) en comparación con los de las células testigo no tratadas con L. casei en los cuatro días de estudio (Figura 3). La mínima cantidad de ON fuede 16.1 a las 96 h, mientras que el valor máximo fuede 26.2 a las 72 h post-exposición a L. casei (Figura 3).
Cuando se compararon los promedios de cuatro días de ON producido por los monocitos en los tres grupos de edad, se observó que hubo una diferencia significativa (p < 0.01) en los valores obtenidos por las células de los becerros de 4-6 meses de edad en comparación con los monocitos de bovinos de ocho y 12 meses de edad (Figura 4) . Los resultados obtenidos en la presente investigación, en bovinos de tres edades, indican que L. casei (bacteria Gram positiva) indujo la producción significativa de ON en cultivos de monocitos expuestos a este lactobacilo en comparación con las células testigo no expuestas, lo que concuerda con Werling et al. (2004) quienes observaron que la producción de ON en cultivos de macrófagos expuestosa distintas bacterias Gram negativas, fue estadísticamente mayor que la producción de ON en células testigo no tratadas. Dichos resultados sugierenque los monocitos fueron activados por de receptores tipo Toll. Al respecto, se sabe que L. casei estimula la respuesta inmunitaria innata a través dela activación de TLRs y la producción de citocinastipo Th1 (Maldonado Galdeano y Perdigón 2006, Vizoso et al. 2009). Es importante resaltar que los monocitos se diferencian en macrófagos y células dendríticas cuando pasan de la circulación a los tejidos y que el bazo es el órgano de almacenamiento de los monocitos (Swirski et al. 2009), por lo 251 que estas células participan de forma activa en los mecanismos inmunitarios innatos. Los resultados de la presente investigación apoyan la idea de quela activación de la inmunidad innata por medio de agonistas de los receptores tipo Toll, como L. casei, también promueve una buena respuesta inmunitaria adaptativa (Bachmann y Jennings 2010) lo cual puede ser aprovechado en el mejoramiento de vacunas (Pashine et al. 2005). Los monocitos debecerros de 4 a 6 meses de edad produjeron una mayor cantidad de ON en comparación con los otros grupos de edad, lo que concuerda con la observación de que los becerros presentan mecanismos inmunitarios innatos superiores a los de los bovinos adultos (Goff et al. 2010).
L. casei indujo la producción de óxido nítrico in vitro en monocitos de bovino de tres grupos diferentes de edad. La mayor producción se observóen becerros de 4-6 meses de edad. Los resultados sugieren que L. casei se podría utilizar en bovinospara estimular la inmunidad innata contra diferentes patógenos, particularmente en animales jóvenes.
Agradecimientos
Este estudio se llevó a cabo con recursos del proyecto 1-1.6-9512932008-P-P.l-l del INIFAP.
LITERATURA CITADA
Adams DO (1979) Macrophages. In: Jakoby W, Pastan I (eds.). Method and Enzymology. Vol. LVIII. Academic Press. New York, pp: 494-506.
Adams JL, Czuprynski CJ (1990) Bacterial lipopolysaccharide induces release of tumor necrosis factor-a from bovine peripheral blood monocytes and alveolar macrophages in vitro. Journal of Leukocyte Biology48: 549-556.
Auffray C, Sieweke MH, Geissmann F (2009) Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annual Review of Immunology 27: 669-692.
Bachmann MF, Jennings CT (2010) Vaccine delivery: a matter of size, geometry, kinetics and molecular patterns. Nature Reviews Immunology 10: 787-796.
Bautista-Garfias CR, Ixta-Rodríguez O, Martínez-Gómez F, López MG, Aguilar BR (2001). Effect of viable or dead Lactobacillus casei organisms administered to mice on resistance against Trichinella spiralisinfection. Parasite 8: S226-S228.
Bautista-Garfias CR, Gómez MB, Aguilar BR, Ixta O, Martínez F, Mosqueda J (2005) The treatment of mice with Lactobacillus casei induces protection against Babesia microti infection. Parasitology Research 97:472-477.
Bautista CR, Alvarez JA, Mosqueda JJ, Falcon A, Ramos JA, Rojas C et al. (2008) Enhancement of the mexican bovine babesiosis vaccine efñcacy by using Lactobacillus casei. Annals of the New YorkAcademy of Sciences 1149: 126-130.
Bautista-Garfias CR, Castañeda R, Álvarez JA, Rojas C, Figueroa JV, Rodríguez A (2012) La vacunación simultánea de bovinos con Lactobacillus casei y la vacuna bivalente contra babesiosis bovina generauna mayor protección contra Babesia bovis y B. bigemina transmitidas por garrapatas en condicionesextremas de campo. Veterinaria México 43: 189-200.
Birmingham JR, Jeska EL (1980) The isolation, long-term cultivation and characterization of bovine peripheral blood monocytes. Immunology 41: 897-814.
Fang FC (1997) Mechanisms of nitric oxide-related antimicrobial activity. Journal of Clinical Investigation 99: 2818-2825.
Goddeeris BM, Baldwin CL, ole-MoiYoi O, Morrison Wl (1986) Improved methods for purification and depletion of monocytes from peripheral blood mononuclear cells. Journal of Immunological Methods89: 165-173.
Goff WL, Bastos RG, Brown WC, Johnson WC, Schneider DA (2010) The bovine spleen: interactions among splenic cell populations in the innate immunologic control of hemoparasitic infections. VeterinaryImmunology and Immunopathology 138: 1-14.
Hume DA, Ross IL, Himes SR, Sasmono RT, Wells CA, Ravasi T (2002) The mononuclear phagocyte System revisited. Journal of Leukocyte Biology 72: 6217627.
Johnson WC, Cluff CW, Wyatt CR, Goff WL (1996) Reactive Oxygen and Nitrogen intermediates and products from polyamine degradation are babesiacidal in vitro. Annals of the New York Academy ofSciences 791: 136-147.
Maldonado Galdeano C, Perdigón G (2006) The probiotic bacterium Lactobacillus casei induces activation of the gut mucosal immune System through innate immunity. Clinical and Vaccine Immunology 13:219-226.
Olivares S (1994) Paquete de diseños experimentales FAUNL, Versión 2.5. Facultad de Agronomía, UANL, Marín, Nuevo León, México.
Pavanelli WR, Nogueira Silva JJ (2010) The Role of Nitric Oxide in immune response against Trypanosoma cruzi infection. The Open Nitric Oxide Journal 2: 1-6.
Pavanelli WR, Salazar Gutierres FR, Nogueira Silva JJ, Conchon Costa I, Noronha Dutra de Menezes MC, de Abreu Oliveira FJ et al. (2010) The effects of nitric oxide on the immune response during giardiasis. Brazilian Journal of Infectious Diseases 14: 606-612.
Pashine A, Valíante NM, Ulmer JB (2005) Targeting the innate immune response with improved vaccine adjuvants. Nature Medicine 11: S63-S68.
Swirski FK, Nhrendorf M, Etzrodt M, Wildgruber M, Cortez-Reta mozo V, Panizzi P et al. (2009) Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science 325: 612-616.
Vega CA, Buening GM, Rodríguez SD. Carson CA. McLaughlin K (1985) Cryopreservation of Babesia bigemina for in vitro cultivation. American Journal of Veterinary Research 46: 421-423.
Vizoso Pinto MG, Rodríguez Gómez M, Seifert S, Watsi S, Holzapfel WH, Franz CMAP (2009) Lactobacilli stimulate the innate immune response and modulate the TLR expression of HT29 intestinal epithelialcells in vitro. International Journal of Food Microbiology 133: 86-93.
Werling D, Hope JC, Howard CJ, Jungi TW (2004).DifferentiaI production of cytokines, reactive oxygen and nitrogen by bovine macrophages and dendritic cells stimulated with Toll-like receptor agonists.Immunology 111: 41-52.