Artículo científico
Dinámica de crecimiento y extracción nutrimental del fruto de litchi (Litchi chinensis sonn) cv. Brewster
Growing dinamyc and nutrient extraction of litchi fruit (Litchi chinensis sonn) cv. Brewster
1Gelacio Alejo-Santiago, 1Gregorio Luna-Esquivel, 2Eduardo Salcedo-Pérez, 3*Rufo Sánchez-Hernández, 1Circe A. Aburto-González.
1Unidad Académica de Agricultura. Universidad Autónoma de Nayarit. Carretera Tepic-Compostela Km. 9, 63780 Xalisco, Nayarit
2Departamento de madera, celulosa y papel. Universidad de Guadalajara.
3División Académica de Ciencias Agropecuarias. Universidad Juárez Autónoma de Tabasco.
*rusaher@hotmail.com
Recibido el 8 de julio de 2013
Aceptado el 22 de agosto de 2014
RESUMEN.
El experimento se desarrolló en un huerto de litchi Cv. Brewster de siete años de edad, el objetivo fue conocer la dinámica de crecimiento del fruto, identificar los momentos críticos de demanda nutrimental y determinar la extracción nutrimental del fruto. Se estableció un experimento completamente al azar en 25 árboles en los que se etiquetaron cuatro frutos por árbol para monitorear el crecimiento desde el día 22 después de la antesis hasta su maduración, se tomaron seis muestras de fruto para cuantificar la extracción de nutrientes, el peso fresco total del fruto, pericarpio, semilla y arilo. Los resultados indican que el patrón de crecimiento del fruto es de carácter sigmoidal simple con un crecimiento intensivo del fruto de 30 d. La acumulación de materia seca y el inicio de formación de arilo ocurre a los 35 d después de la antesis; la composición del fruto al momento de la cosecha es de 22 % de pericarpio, 30 % de semilla y 48 % de mesocarpio. La extracción nutrimental en kg por tonelada de fruto fresco es N 2.8, P2O5 1.1, K2
Palabras clave: Nutrición, materia seca, fertilización, producción.
ABSTRACT.
The experiment was conducted in a 7 years oíd Brewster litchi orchard, the aim was to determine the dynamic of fruit growth, identify the critical moments of nutrient demand and determine fruit nutrient extraction. A completely randomized experiment was held in 25 trees in which 4 fruits were labeled to monitor from day 22 after anthesis to maturity; six samples of fruit were taken to quantify fruit nutrient extraction, total weight of fresh fruit, pericarp, seed and aril. Results indícate that the pattern of fruit growth is simple sigmoidal character with an intensive fruit growth of 30 days. Dry matter accumulation and aryl formation begin at 35 d after anthesis; the composition of the fruit at harvest time is 22 % pericarp, 30 % seed and 48 % of mesocarp. The nutrient extraction in kg per ton of fresh fruit is N 2.8, P205 1.1, K2
Key words: Nutrition, dry matter, fertilization, production.
INTRODUCCIÓN
El litchi (Litchi chinensis Sonn.) es un frutal introducido recientemente en el mercado internacional, que ha registrado un continuo crecimiento de su demanda, lo que ha generado un déficit en la producción (Singh et al. 2012). A nivel mundial los principales productores son China, Israel, Australia, Tailandia, India, Vietnam, algunas partes de África, México, Centro y Sudamérica; estimándose que el 95 % del área cultivada se encuentra en Asia. En el 2002, en esa área del mundo se tenían establecidas 600,000 ha, de las cuales el 60 % se establecieron en los últimos diez años, y se observaba un rápido crecimiento en la superficie cultivada (Evans y Degner 2005). En el ámbito nacional, se tiene registrada una superficie de 3 571.4 ha en las que se producen 18 062.4 t (SIAP 2012), y son al menos en 14 estados de la república mexicana donde se tienen condiciones adecuadas de precipitación, vientos y temperaturas que permiten el cultivo de este frutal (De la Garza 2004); De acuerdo con Singh et al. (2012) el litchi es un cultivo que alcanza altos precios en el mercado internacional, por lo que representa una opción viable para diversificar la agricultura de aquellos países que ofrecen las condiciones agroclimatológicas para su cultivo, entre ellos México. No obstante, Menzel y Simpson (1987) mencionan que el crecimiento en la superficie cultivada ha sido restringido debido a múltiples factores, dentro de los más significativos se encuentra la baja producción por árbol que se obtiene, así como la presencia de fenómenos fisiológicos como la alternancia; la cual se atribuye a diversas causas, entre las que se encuentran el escaso conocimiento sobre la fenología floral, plagas, enfermedades y demanda nutrimental. Esta baja producción provoca un incremento en los costos de producción y disminuyen la rentabilidad del cultivo (Osuna et al. 2008, Wilkie et al. 2008).
Algunas de las formas de atenuar el efecto de estos factores restrictivos, son un adecuado manejo, y el empleo de algunas técnicas basadas en los procesos fisiológicos relacionados con la fenología, iniciación floral, floración, caída de frutos y la extracción nutrimental del fruto en sus distintas etapas de crecimiento (Ying y Davenport 2004). El fenómeno de alternancia es uno de los problemas que presenta el cultivo del litchi, el cual se asocia con factores genéticos y climáticos, particularmente con el déficit de temperaturas bajas en otoño-invierno, necesarias en los procesos de floración y amarre de fruto (García y Martins 2006). Otra de las explicaciones que se ha dado a este fenómeno, es que puede ser causado por un desbalance nutrimental, tal como sucede en otros frutales como los cítricos, donde la floración, el crecimiento y la producción de fruta utilizan grandes cantidades de carbohidratos, por lo que después de una alta producción, se registra un déficit de nutrientes en el ciclo siguiente, lo que da lugar a un año de baja producción; que puede ser la principal causa de la reducción en la acumulación de carbohidratos.
Lo que da como resultado una baja floración y un mayor desarrollo vegetativo (Ruíz 2001). De acuerdo con Taub y Lerdau (2000), en la producción de carbohidratos confluyen diversos elementos entre los que se encuentran los nutrimentos esenciales para las plantas, particularmente existe una relación directa entre la concentración de nitrógeno (N) en hojas y la fotosíntesis neta. En cítricos, se ha demostrado que el amarre de frutos pende también de la disponibilidad de carbohidratos (Iglesiaset al. 2003), y una vez que se da el amarre, el crecimiento activo de estos depende de la disponibilidad de agua, fotoasimilados y minerales (Iglesias et al. 2007). En arboles de mango (Manguifera indica) la capacidad fotosintética tiene una estrecha correlación no lineal con la concentración de N en las hojas; esto se debe a que entre el 40 y 50 % del N foliar, es asignado a las funciones de carboxilación y portación de electrones, lo que se traduce en un incremento en la tasa fotosintética (Urban et al. 2003).
Por su parte, el fósforo (P) es un elemento móvil en la planta y se incorpora de forma rápida al metabolismo (Schachtman et al. 1998). En primer lugar se producen azúcares y alcoholes fosforilados como productos intermedios, así como los fosfolípidos que son componentes básicos de las membranas celulares (Domínguez 1997). Antes de ser metabolizados, los azúcares deben ser fosforilados, para lo cual se requiere ATP, la disminución en el nivel de ATP que caracteriza a plantas cultivadas en deficiencia de P puede limitar el metabolismo de los azúcares (Theodorou et al. 1991). El P es un participante básico en casi todos los procesos de crecimiento y síntesis de compuestos constituyentes, en condiciones de deficiencia se retrasa la madurez del fruto, además de requerirse para la formación de semilla (Navarro y Gines 2000).
Otro nutrimento ampliamente estudiado es el potasio (K), el cual es vital en la fotosíntesis (Szczerba et al. 2009), cuando hay deficiencias, ésta se reduce e incrementa la respiración, por lo que disminuye la acumulación de carbohidratos y ocasiona un efecto adverso en el crecimiento y producción de la planta (Ooterhuis et al. 2014). El K está presente en todo el tejido vegetal; es un componente importante de los suelos fértiles y se requiere en grandes cantidades por las plantas (Singh et al. 2012).
La nutrición es un factor considerado como controlable dentro de un sistema de producción, pero no existen datos específicos en cuanto a la extracción nutrimental del litchi que permitan precisar la dosis óptima de fertilización; por lo que no se puede establecer una nutrición balanceada que favorezca la obtención de rendimientos altos y constantes. Por ello es necesario conocer la dinámica de crecimiento y las tendencias de extracción nutrimental del fruto, que permita establecer programas de auxilio en aplicación de productos fertilizantes e identificar los momentos críticos de demanda (Diczbalis et al. 2010). Al respecto Galán y Menini (1987) mencionan que por cada tonelada de fruto fresco de litchi, se extrae 2.2 kg de N, 2.2kg de P2O5, 6.6 kg de K2O, 1.6 kg de CaO y 1.1 kg de MgO.
Tomando en cuenta que existe una alta demanda internacional de fruta de litchi y que México es un país que tiene condiciones climáticas para producir y satisfacer parte de esta demanda, pero que no cuenta con suficiente información técnica y científica referente a este cultivo, los objetivos del presente trabajo fueron determinar la extracción nutrimental de los frutos e identificar los momentos críticos de la demanda en árboles de litchi Brewster.
MATERIALES Y MÉTODOS
Caracterización del sitio de estudio
El trabajo se realizó en un huerto de litchi ’Brewster’, localizado en el ejido Barranca Blanca, municipio de Tepic, Nayarit, con árboles de siete años de edad, sembrados a una densidad de 204 árboles ha-1. El clima corresponde a un (A)e (W2) a (i), que se define como un clima semicálido (subtropical subhúmedo), el más cálido de los templados (c). El régimen pluviométrico es mayor de 1 300 mm anuales; el mes de máxima precipitación es jJulio con 370 a 380 mm, mientras que el de menor precipitación es mayo, con menos de 30 mm. La temperatura media anual varía de 20a 29 °C, siendo los meses de junio a septiembre los más cálidos, con una temperatura media de 23 a 24 °C, y de diciembre a enero, los más fríos, con temperaturas promedio de 16 a 17 °C (García 2004).
Análisis físicos y químicos de suelo
En los cuatro puntos cardinales del tronco de cada uno de los árboles seleccionados, se colectaron submuestras de suelo a una profundidad de 0 a 40 cm para formar una muestra compuesta, dichas muestras fueron a la sombra, para posteriormente molerlas y tamizarlas, para determinar pH, conductividad eléctrica, textura, materia orgánica, fósforo, bases intercambiables (Ca2+, Mg2+ y K+). Todos los análisis físicos y químicos se realizaron de acuerdo con los recomendado por Chapman y Pratt (1979), y la Norma Oficial Mexicana PROY-NOM-021-RECNAT (DOF 2000), en el laboratorio de Suelo, Agua y Planta, de la Unidad Académica de Agricultura de la Universidad Autónoma de Nayarit.
Planeación del experimento y manejo del huerto
Se seleccionaron 25 arboles al azar; homogéneos en edad, tamaño, vigor, diámetro del tallo, sanidad y carga de fruto; cada árbol se consideró como una unidad experimental. El manejo agronómico consistió en dos aplicaciones anuales de glifosato (3.5 L ha-1) para el control de malezas y se realizaron cuatro fertilizaciones al año en los meses de febrero, marzo, abril y mayo de 2012, con un kg de la mezcla 18-46-00 NPK y 250 g de sulfato de potasio por árbol. La aplicación se realizó al suelo, a una distancia de 0.20 m de la base del tallo. Se regaron los árboles una vez por semana, con un gasto aproximado de 300 I de agua por planta, distribuido en un radio de 0.75 m, para llevar el suelo a capacidad de campo en una profundidad de 0.4 m, estos riegos se efectuaron al inició de brotación de yemas reproductivas y se continuaron hasta la maduración de fruto. Durante la floración se realizó una aplicación de Oxicloruro de cobre (Cupravit) (2 g L-1) como tratamiento preventivo para el control de enfermedades fungosas.
Determinación de la dinámica del crecimiento del fruto
Cuando los árboles presentaban frutos de aproximadamente 0.5 cm de longitud; en cada árbol se seleccionaron y etiquetaron cuatro frutos, uno por cada punto cardinal, por lo que en total, se monitorearon 100 frutos. A dichos frutos se les registró la longitud cada cuatro días, hasta que estos alcanzaron tamaños longitudinales promedios de 0.9, 1.3, 1.7, 2.4, 2.8, 3.3 (inicio de maduración) y 3.3 cm (fruto maduro). Estos frutos etiquetados permanecieron en los arboles hasta su madurez, y fueron utilizados como indicadores para la colecta de otros frutos de tamaño similar, en los que se realizaron los análisis químicos para determinar la extracción nutrimental.
Durante el periodo de crecimiento, se realizaron siete muéstreos. En cada uno de ellos, se seleccionaron cuatro frutos por árbol, mismos que en el momento del muestreo tuvieron una longitud de 0.9, 1.3, 1.7, 2.4, 2.8, 3.3 (inicio de maduración) y 3.3 cm (fruto maduro); dimensiones que fueron similares a la de los frutos indicadores. Los muéstreos del uno al seis fueron utilizados para determinar la dinámica de extracción de nutrimentos, mientras que el muestreo siete fue utilizado para cuantificar la concentración nutrimental en cada uno de los componentes del fruto en el momento de la cosecha.
Obtención y procesamiento de muestras
Los frutos colectados en cada muestreo, se separaron en pericarpio, semilla y mesocarpio para determinar su peso fresco, excepto en los muéstreos uno y dos, donde solamente fue posible cortar los frutos completos en trozos pequeños. Los diferentes componentes del fruto fueron colocados en platos de aluminio, para secarlos en una estufa de aire forzado Lumistell® HTP-42 a 65 °C hasta peso constante (Alcántar y Sandoval 1999). Las muestras secas, se molieron en un molino de cuchillas de acero inoxidable Thomas Willey Mili modelo ED-5®, de las cuales se tomaron 0.5 g del tejido para determinar el N-total por el método de Kjeldal; Ca2+, Mg2+, Zn2+, Mn y Fe2+, por espectrofotometría de absorción atómica (Alcántar y Sandoval 1999); P por molibdato de amonio y K+ por Flamometría (Chapman y Pratt 1979).
Procesamiento estadístico de los datos
La información obtenida fue procesada mediante análisis de varianza y Prueba de Tukey (α ≤ 0.05) mediante el programa Statistical Analysis System (SAS Institute Inc. 2009). Considerando como tratamientos los momentos de muestreo y como variables dependientes: concentración nutrimental, proporción de mesocarpio, cáscara y semilla. El modelo de crecimiento de fruto se obtuvo mediante un análisis de regresión no lineal, con la utilización del complemento XLSTAT (Ver. 7.5.2) de Excel 2003.
RESULTADOS
Descripción de la fertilidad del suelo
Los resultados de los análisis iniciales de suelo registraron contenido de materia orgánica de 2.9 %, conductividad eléctrica de 0.05 dS m-1, pH de 6.8; concentración de P de 6.29; 109 mg kg-1 de K, 3755 mg kg-1 de Ca, y 197 mg kg-1 de Mg; 3 755 mg kg-1 de Ca, y 197 mg kg-1 de Mg; composición granulométrica de 60 % de arena, 28 % de limo y 12 % de arcilla, que lo clasifica como de textura arenosa. Los valores de las variables edáficas se consideraron como óptimas para el buen desarrollo del cultivo, aunque hay nutrientes que se encontraron en bajas concentraciones como es el caso del P(< 15 mg kg-1) y K.
Dinámica de crecimiento de fruto
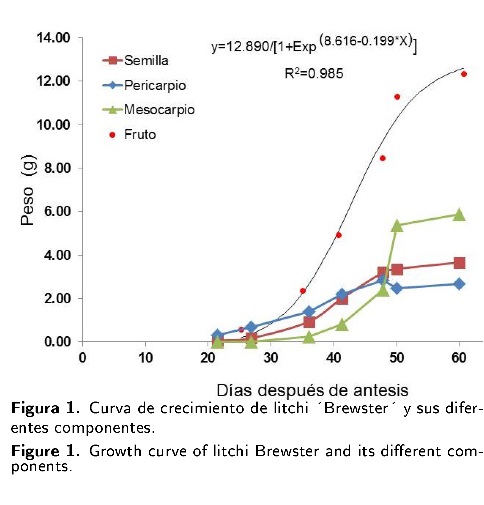
La curva de crecimiento de fruto indica un comportamiento sigmoidal, por lo que se requirieron alrededor de 26 d después de la antesis (dda) para iniciar un crecimiento acelerado del fruto, además se puede observar que a partir de ese momento transcurren 30 dda hasta que el fruto alcanza su máximo peso, para luego iniciar la etapa de maduración del fruto; se observa que el mesocarpio comienza a ganar peso a los 36 dda, y tiene una duración aproximada de 20 d, para luego iniciar la maduración del fruto (Figura 1).
Peso fresco de fruto y de sus partes
Se observó que conforme avanza el periodo de crecimiento del fruto, el peso del pericarpio disminuye. Caso contrario a lo que sucede con el mesocarpio, en donde, el inicio de acumulación de peso comienza a partir de los 36 dda y alcanza su porcentaje máximo a los 50 dda, registrando hasta un 48 % de peso con respecto al peso fresco total del fruto. La semilla del fruto fue el componente que tuvo un menor rango de variación, al pasar de 30.01 % a 39.89 %, con respecto al peso total del fruto. En el fruto maduro, la distribución de materia fresca fue 48.03 % para mesocarpio, 30 % semilla y 21.97 % pericarpio (Tabla 1).
Concentración nutrimental en pericarpio
La concentración de nutrientes en pericarpio, muestra que algunos nutrimentos como el N, P, K, disminuyen conforme el fruto crece; para el caso de Ca se elevó la concentración en la etapa final de crecimiento del fruto, esta situación es producto de la concentración de este nutrimento en el suelo, ya que según el análisis de suelo, el huerto posee alta concentración de Ca (> 3000 mg kg-1), por lo que se considera como una condición de suficiencia para este nutrimento. En cuanto al Mg, Mn, Fe y Zn, no se presentaron diferencias significativas a través del periodo de crecimiento; mientras que el Cu mostró mayor contenido a los 36 y 41 dda, esto pudo deberse a la aplicación de productos fungicidas a base de Cu en el huerto (Tabla 2).
Concentración nutrimental en semilla
Los nutrimentos mayormente concentrados en semilla fueron K y N, mientras que en cuanto a micronutrientes fueron el Cu, Fe y Zn, en los que no se observaron variaciones en su concentración durante el periodo de muestreo (Tabla 3). Los valores encontrados de concentración de micronutrientes indican que conforme va creciendo el fruto hay una demanda constante de estos nutrimentos.
Concentración nutrimental en mesocarpio
Los nutrimentos de mayor concentración fueron el K y el N, por lo que es muy importante realizar aplicaciones de auxilio de este nutriente vía foliar durante el desarrollo del fruto, lo que además mejora su concentración de azucares e incrementa su calidad; los resultados encontrados muestran que conforme creció el fruto, hubo nutrientes que disminuyeron en concentración (Tabla 4).
Extracción nutrimental de fruto
El contenido de humedad en los componentes fue de 60 % en pericarpio y 54.5 % en semilla. La cantidad de nutriente extraído por tonelada de fruto fresco fue: 2.6 kg de N, 1.1 kg de P2O5, 3.4 kg de K2O, 0.94 kg de CaO, 0.12 kg de MgO, 0.06 de Cu, 5.46 g, de Fe 3.84 g de Mn y 15.48 g de Zn.
DISCUSIÓN
La dinámica de crecimiento del fruto del litchi fue del tipo sigmoidal; lo que significa que hay una etapa en la que se alcanza un mayor punto de crecimiento; a partir de dicho punto, el crecimiento es más ligero y constante (Vieira et al. 1996). En el caso de este fruto, este punto se registró a los 50 dda. Al respecto, Salomáo et al. (2006) reportaron un patrón de crecimiento sigmoidal en litchi "Bengal" que es similar a los registrados en el presente trabajo, aunque los datos de distribución porcentual de pericarpio, semilla y mesocarpio, sobre todo en la etapa de fruto maduro fueron diferentes; ellos mencionaron que el peso del fruto se distribuye en 37.8 % para pericarpio, 32 % a semilla y 30.2 % al mesocarpio, por lo que de acuerdo a los resultados de la presente investigación, existe una diferencia de casi 20 % en el mesocarpio entre "Brewster" y "Bengal", ya que "Brewster" puede llegar a registrar hasta 48 % con respecto al peso total del fruto.
En el presente trabajo se observó que la proporción de los componente del fruto (pericarpio, semilla y mesocarpio) con respecto al peso total, siguió cinéticas distintas a partir de los 36 dda; en el caso de la semilla, se observó una baja permanentemente hasta estabilizarse a los 50 dda; en ese momento la proporción porcentual de este componente fue de 21.94 % con respecto al peso total del fruto. Por su parte, la semilla fue una estructura del fruto que mantuvo un incremento hasta a los 40 dda. A partir de ese momento hubo una disminución proporcional, hasta estabilizarse a los 50 dda; la proporción porcentual de la semilla con respecto al peso total del fruto fue de 30 %. El mesocarpio o arilo es una estructura del fruto que mantiene un crecimiento continuo, y puede llegar a crecer ligeramente 10 d más con respecto a los demás componentes. De acuerdo con Lira (2007), el crecimiento se define como el incremento natural en tamaño, donde intervienen diversos factores, entre los que destaca una mayor demanda de nutrimentos, por lo que en esta etapa fisiológica del fruto, se genera un incremento en dicha demanda. En este sentido, se observó que precisamente fue entre 36 y 41 dda, en donde se observaron las mayores concentraciones de los elementos evaluados; aunque la tendencia en las concentraciones nutrimentales fue a la baja a partir del 41 dda, observándose un repunte en los contenidos de N, P, Ca, Zn y Mn hacia etapas de mayor madurez del fruto (50 dda), sin que estos contenidos lleguen a los niveles registrados durante los inicios de la etapa intensa del crecimiento. De acuerdo con Greenwood (1983), la demanda de nutrimentos por un cultivo está dada por su producción de biomasa y la concentración del nutrimento en la planta. Esto explica por qué la mayor concentración nutrimental ocurrió entre los 36 y 41 dda, ya que es el periodo en donde el fruto experimentó un crecimiento más acelerado. Al estabilizarse el crecimiento del fruto a los 41 dda, la concentración de la mayoría de los nutrimentos disminuyó. Sin embargo, a diferencia del pericarpio y la semilla que detuvieron su crecimiento a partir del 41 dda; el mesocarpio aún continuó creciendo, esa puede ser la causa que generó de nueva cuenta un incremento en la demanda nutrimental, misma que se reflejó en el repunte en la concentración de los nutrientes N, P, Ca, Zn y Mn, hacia la etapa final de crecimiento del fruto. Al respecto Molina et al. (1993) y Sancho (1999) señalaron que el conocimiento de las épocas de mayor absorción nutrimental, puede ayudar a definir un adecuado programa de fertilización, en el cual se considere la dosis de fertilizante y la época adecuada para hacer las aplicaciones. Los resultados de este estudio indican que 80 % de la concentración nutrimental en fruto, ocurre durante los primeros estadios de crecimiento. Por ello, dependiendo de las fuentes fertilizantes que se utilicen, se debe considerar que la mayor tasa de disponibilidad de nutrimentos debe darse justamente en la etapa inicial de la fructificación. Trabajos experimentales realizados por Maldonato et al. (2012), en los que se evaluó el efecto de la fertilización en arboles de litchi, indicaron que la producción de fruto por árbol y el contenido de clorofila se incrementa en un 25 % al portar la dosis 50-130-50 (N, P2O5 y K2); y que la mayor concentración nutrimental en el área foliar es del N (1.5 %), seguido del K (1.1 %) y el P (0.22 %), con niveles críticos variables para cada elemento; para el caso del N es de 1.5 a 1.8 %, para el P de 0.14 a 0.22 % y el K de 0.7 a 1.1 %. Esas concentraciones nutrimentales en el área foliar que reportaron los autores mencionados, contrastan con las concentraciones obtenidas en los diferentes componentes del fruto; ya que en el presente trabajo, los resultados indicaron que con base en la concentración nutrimental, los macronutrientes más demandados son K, N, Ca, P y Mg; mientras que para el caso de los micronutrientes son Zn, Fe, Mn y Cu. Se observó que esta concentración de K es mayor en las primeras etapas de crecimiento del fruto, y particularmente en el pericarpio, aunque hacia la parte final de crecimiento del fruto, la mayor concentración ocurre en semilla y mesocarpio. Estos resultados son congruentes con los presentados por Galán y Menini (1987), los cuales refieren que en el cultivo de litchi, el nutrimento de mayor demanda es el K, y que se requiere de 6.6 kg de K20 para producir una tonelada de fruto fresco. Según Domínguez (1997), el K es responsable de la función de transporte de azucares hacia los órganos de demanda. Por ello, si se considera que los huertos establecidos en el área de estudio son deficientes en K, tal a como lo revelan los estudios previos, es necesario implementar un programa de fertilización que considere la restitución de los nutrimentos que se extraen en mayor cantidad, tal como sucede con el K. Hipotéticamente, el fenómeno de la alternancia obedece entre otros factores déficit nutrimentales provocado por una alta extracción de nutrimentos durante la cosecha, que si no se reponen para el ciclo siguiente ocurre una baja en el rendimiento. Por ello, los programas de fertilización en litchi deben de poner especial atención a nutrimentos de alta extracción como el K. Otras investigaciones realizadas en Sapindáceas como el longan (Dimocarpus brigán), han reportado una buena respuesta de la producción a las aplicaciones de clorato de potasio (Evans y Nalampang 2009).
La dinámica de crecimiento del fruto del litchi fue del tipo sigmoidal; lo que significa que hay una etapa en la que se alcanza un mayor punto de crecimiento; a partir de dicho punto, el crecimiento es más ligero y constante (Vieira et al. 1996). En el caso de este fruto, este punto se registró a los 50 dda. Al respecto, Salomáo et al. (2006) reportaron un patrón de crecimiento sigmoidal en litchi 'Bengal' que es similar a los registrados en el presente trabajo, aunque los datos de distribución porcentual de pericarpio, semilla y mesocarpio, sobre todo en la etapa de fruto maduro fueron diferentes; ellos mencionaron que el peso del fruto se distribuye en 37.8 % para pericarpio, 32 % a semilla y 30.2 % al mesocarpio, por lo que de acuerdo a los resultados de la presente investigación, existe una diferencia de casi 20 % en el mesocarpio entre "Brewster" y "Bengal", ya que "Brewster" puede llegar a registrar hasta 48 % con respecto al peso total del fruto. En el presente trabajo se observó que la proporción de los componente del fruto (pericarpio, semilla y mesocarpio) con respecto al peso total, siguió cinéticas distintas a partir de los día 36 dda; en el caso de la semilla, se observó una baja permanentemente hasta estabilizarse a los 50 dda; en ese momento la proporción porcentual de este componente fue de 21.94 % con respecto al peso total del fruto. Por su parte, la semilla fue una estructura del fruto que mantuvo un incremento hasta a los 40 dda. A partir de ese momento hubo una disminución proporcional, hasta estabilizarse a los 50 dda; la proporción porcentual de la semilla con respecto al peso total del fruto fue de 30 %. El mesocarpio o arilo es una estructura del fruto que mantiene un crecimiento continuo, y puede llegar a crecer ligeramente 10 d más con respecto a los demás componentes. De acuerdo con Lira (2007), el crecimiento se define como el incremento natural en tamaño, donde intervienen diversos factores, entre los que destaca una mayor demanda de nutrimentos, por lo que en esta etapa fisiológica del fruto, se genera un incremento en dicha demanda. En este sentido, se observó que precisamente fue entre 36 y 41 dda, en donde se observaron las mayores concentraciones de los elementos evaluados; aunque la tendencia en las concentraciones nutrimentales fue a la baja a partir de los 41 dda, observándose un repunte en los contenidos de N, P, Ca, Zn y Mn hacia etapas de mayor madurez del fruto (50 dda), sin que estos contenidos lleguen a los niveles registrados durante los inicios de la etapa intensa del crecimiento. De acuerdo con Greenwood (1983), la demanda de nutrimentos por un cultivo está dada por su producción de biomasa y la concentración del nutrimento en la planta. Esto explica por qué la mayor concentración nutrimental ocurrió entre los 36 y 41dda, ya que es el periodo en donde el fruto experimentó un crecimiento más acelerado. Al estabilizarse el crecimiento del fruto a los 41 dda, la concentración de la mayoría de los nutrimentos disminuyó. Sin embargo, a diferencia del pericarpio y la semilla que detuvieron su crecimiento a partir del 41 dda; el mesocarpio aún continuó creciendo, esa puede ser la causa que generó de nueva cuenta un incremento en la demanda nutrimental, misma que se reflejó en el repunte en la concentración de los nutrientes N, P, Ca, Zn y Mn, hacia la etapa final de crecimiento del fruto. Al respecto Molina et al. (1993) y Sancho (1999) señalaron que el conocimiento de las épocas de mayor absorción nutrimental, puede ayudar a definir un adecuado programa de fertilización, en el cual se considere la dosis de fertilizante y la época adecuada para hacer las aplicaciones. Los resultados de este estudio indican que 80 % de la concentración nutrimental en fruto, ocurre durante los primeros estadios de crecimiento. Por ello, dependiendo de las fuentes fertilizantes que se utilicen, se debe considerar que la mayor tasa de disponibilidad de nutrimentos debe darse justamente en la etapa inicial de la fructificación. Trabajos experimentales realizados por Maldonato et al. (2012), en los que se evaluó el efecto de la fertilización en arboles de litchi, indicaron que la producción de fruto por árbol y el contenido de clorofila se incrementa en un 25 % al aportar la dosis 50-130-50 (N, P2O5 y K2O); y que la mayor concentración nutrimental en el área foliar es del N (1.5 %), seguido del K (1.1 %) y el P (0.22 %), con niveles críticos variables para cada elemento; para el caso del N es de 1.5 a 1.8 %, para el P de 0.14 a 0.22 % y el K de 0.7 a 1.1 %. Esas concentraciones nutrimentales en el área foliar que reportaron los autores mencionados, contrastan con las concentraciones obtenidas en los diferentes componentes del fruto; ya que en el presente trabajo, los resultados indicaron que con base en la concentración nutrimental, los macronutrientes más demandados son K, N, Ca, P y Mg; mientras que para el caso de los micronutrientes son Zn, Fe, Mn y Cu. Se observó que esta concentración de K es mayor en las primeras etapas de crecimiento del fruto, y particularmente en el pericarpio, aunque hacia la parte final de crecimiento del fruto, la mayor concentración ocurre en semilla y mesocarpio. Estos resultados son congruentes con los presentados por Galán y Menini (1987), los cuales refieren que en el cultivo de litchi, el nutrimento de mayor demanda es el K, y que se requiere de 6.6 kg de K20 para producir una tonelada de fruto fresco. Según Domínguez (1997), el K es responsable de la función de transporte de azucares hacia los órganos de demanda. Por ello, si se considera que los huertos establecidos en el área de estudio son deficientes en K, tal a como lo revelan los estudios previos, es necesario implementar un programa de fertilización que considere la restitución de los nutrimentos que se extraen en mayor cantidad, tal como sucede con el K. Hipotéticamente, el fenómeno de la alternancia obedece entre otros factores déficit nutrimentales provocado por una alta extracción de nutrimentos durante la cosecha, que si no se reponen para el ciclo siguiente ocurre una baja en el rendimiento. Por ello, los programas de fertilización en litchi deben de poner especial atención a nutrimentos de alta extracción como el K. Otras investigaciones realizadas en Sapindáceas como el longan (Dimocarpus brigán), han reportado una buena respuesta de la producción a las aplicaciones de clorato de potasio (Evans y Nalampang 2009).
Por otra parte, ya se mencionó que el N fue el segundo nutrimento de mayor concentración en el fruto del litchi, particularmente en la semilla donde en promedio puede llegar a acumular 7.14 g kg-1; según Pathak y Mitra (2010) el N juega un papel importante en el crecimiento y desarrollo del área foliar, por lo que el efecto sobre el crecimiento del fruto sería indirecto, mientras que el suministro de K tiene un efecto más directo y de mayor relevancia en el rendimiento y calidad del fruto. Al respecto, Khaosumain et al. (2013) señalaron que un incremento en la aplicación de N tiende a incrementar el área foliar, peso fresco y seco de las hojas; sin embargo, cuando se aplican de 320 y 640 g de N por árbol, disminuye el porcentaje de floración y no se observa un efecto sobre el número y peso de frutos por inflorescencia, en tanto que el tamaño del fruto, el grosor de pericarpio y cantidad de mesocarpio, son más bajos cuando se incrementan dichos niveles de N. Los resultados obtenidos en el presente trabajo dejan en claro que la fertilización debe de responder a los requerimientos de nutrimentos en sus diferentes etapas fenológicas, en el caso del litchi por ser un fruto que demanda cantidades de 9.11 y 7.14 g kg -1 de K y N respectivamente, es necesario realizar investigaciones para evaluar la respuesta de la calidad y el rendimiento a la aplicación de ambos nutrimentos.
CONCLUSIONES
El patrón de crecimiento del fruto es sigmoidal simple, observándose que el crecimiento intensivo inicia a los 30 dda, cuando el fruto alcanza 2.5 gramos. Los momentos críticos en acumulación de materia seca e inicio de formación de mesocarpio tiene lugar a los 35 d después de antesis. Se observó que para el caso de este cultivar y bajo las condiciones del presente experimento, la partición de los componentes del fruto, se distribuyen de la son, el K con 3.4 kg t-1 de frutos cosechados, le siguiente manera: pericarpio 22 %, semilla 30 % y mesocarpio 48 %. Los nutrimentos más extraídos son, el K con 3.4 kg t-1 de frutos cosechados, le siguen N, P, Ca y Mg, con 2.8, 1.1, 0.94 y 0.12 kg h-1, respectivamente.
LITERATURA CITADA
Alcántar GG, Sandoval VM (1999) Manual de análisis químico de tejido vegetal. Publicación Especial 10. Sociedad Mexicana de la Ciencia del Suelo, A. C. Chapingo, México. 155 p.
Camilo, AP., Palladini L A (2000) Efeito de diferentes volumes de calda no raleio químico de frutos da macieira Gala. Pesquisa Agropecuária Brasileira 35: 2191-2195.
Chapman HD, Pratt PF (1979) Métodos de análisis para suelos, plantas y agua. Trillas. México, D.F. 195 P-
De la Garza NA (2004) El litchi: alternativa para la diversificación frutícola en la huasteca potosina. Folleto técnico No. 2. Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. México. 40 p.
Diario Oficial de la Federación (DOF) (2000) Proyecto de Norma Oficial Mexicana PROY-NOM-021-RECNAT-2000, que establece las especificaciones de fertilidad, salinidad y clasificación de suelos. Estudios, muéstreos y análisis. Órgano del Gobierno Constitucional de los Estados Unidos Mexicanos. Tomo DLXV-12:6-74.
Diczbalis Y, Nicholls B, Groves I, Lake K (2010) Sapindaceae production and research in Australia. Acta Horticulturae. 863: 49-58
Domínguez VA (1997) Tratado de fertilización. Ediciones Mundi-prensa. México. 613 p.
Evans EA, Nalampang S (2009) Importación de Frutas Tropicales de Tailandia. EDIS FE803 (español versión FE719). Food and Resource Economics Departamento, Florida Cooperative. Extensión Service, Institute of Food and Agricultural Sciences, University of Florida, Gainesville, FL. https://edis.ifas.ufl.edu/pdffiles/FE/FE80300.pdf. Fecha de consulta 15 de febrero de 2014.
Evans EA, Degner RL (2005) Recent developments in world production and trade of Lychee (Litchi chinensis): Implications for Florida growers. Procedings of the Florida State Horticultural Society 118: 247-249
Galán SV, Menini UG (1987) El litchi y su cultivo. Estudio FAO, Producción y Protección Vegetal 83. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Roma, Italia. 205 p.
García E (2004) Modificaciones al sistema de clasificación climática de Kóppen. 5a ed. Serie Libros. Instituto de Geografía. Universidad Nacional Autónoma de México. México, D.F. 90 p.
García PE, Martins GAB (2006) Flowering and fruiting of lychee trees in response to girdling of branches. Revista Brasileira de Fruticultura 28: 14-17.
Greenwood DJ (1983) Quantitative theory and the control of soil fertility. New Phytol. 94: 1-18. Iglesias DJ, Cercos M, Colmenero-Flores JM, Naranjo MA, Ríos G, Carrera E, Ruiz-Rivero O, Lliso I Morillon R, Tadeo FR, Talón M (2007) Physiology of citrus fruiting. Brazilian Journal of Plant Physiology 19: 333-362.
Iglesias DJ, Francisco RT, Primo-Millo E,Talón M (2003) Fruit set dependence on carbohydrate availabilitty in citrus tres. Tree Physiology 23: 199-204.
Khaosumain Y, Sritontip C, Changjeraja S (2013) Effects of different nitrogen fertilization doses on growth, leaf nutrient concentration, flowering and fruit quality in off-season longan. Acta Horticulturae 984: 271-274
Lira S RH (2007) Fisiología vegetal. Trillas, Universidad Agraria Antonio Narro. México. 237 p.
Maldonado PR, Trinidad S A, Téliz O D, Velasco V VA, Volke H VH (2012) Respuesta del litchi (Litchi chinensis Sonn.) a la fertilización con NPK en el norte de Oaxaca, México. Revista Fitotecnia Mexicana 35: 251-258.
Menzel CM, Simpson DR (1987) Lychee Nutrition: A review. Scientia Horticulture 31: 194-224.
Molina E, Salas R, Castro A (1993) Curva de crecimiento y absorción de nutrimentos en fresa (Fragaria x ananassa Duch. cv. Chandler) en Alajuela. Agronomía Costarricense 17: 67-73.
Navarro BS, Gines NG (2000) Química agrícola. Ed. Mundi-Prensa. España. 488 p.
Oosterhuis DM, Loka DA, Kawakami EM, Pettigrew WT (2014) The physiology of potassium in crop pro-duction. Advances in Agronomy 126: 203-233.
Osuna ET, Valenzuela RG, Muy RMD, Gardea BAA, Villareal RM (2008) Expresión del sexo y anatomía floral del litchi (Litchi chinensis Sonn.). Revista Fitotecnia Mexicana 31: 51-56.
Pathak PK, Mitra SK (2010) Rate and time of potassium fertilization incluence yield and quality of litchi. Acta Hosticultura 836: 235-242
Ruiz R, García-Luis A, Monerri C, Guardiola JL (2001) Carbohydrate Availability in Relation to fruitlet Abscission in Citrus. Annals of Botany 87: 805-812.
Salomao LCC, De-Siqueira DL, Pereira MEC (2006) Desenvolvimento do fruto da lichieira (Litchi chinensis sonn.) bengal. Revista Brasileria de Fruticultura 8: 11-13.
Sancho, VH (1999) Curvas de absorción de nutrientes: Importancia y usos en los programas de fertilización. Instituto de la Potasa y el Fósforo. Informaciones Agronómicas 36: 11-13
SAS Institute (2009) SAS/STAT® 9.2 Userss Guide Release. Cary, NC: SAS Institute Inc. USA. 60 p.
Schachtman DP, Reid JR, Ayling SM (1998). Phosphorus uptake by plants: From soil to cell. Plant Physiology 116: 447-453.
Schachtman DP, Reid JR, Ayling SM (1998). Phosphorus uptake by plants: From soil to cell. Plant Physiology 116: 447-453.
SIAP (2012) Servicio de Información Agroalimentaria y Pesquera, http://www.siap.gob.mx. Fecha de consulta 12 de noviembre de 2013.
Singh G, Nath V, Pandey SD, Ray PK, Singh HS (2012) The litchi. Food and Agriculture Organization of the United Nations. New Delhi, India. 219 p.
Szczerba MW, Britto DT, Kronzucker HJ (2009) K+ transport in plants: Physiology and molecular biology. Journal of plant physiology 166: 447-466.
Taub DR, Lerdau MT (2000) Relationship between leaf nitrogen and photosynthetic rate for three NAD-ME and three NADP-ME C4 grasses. American Journal of Botany 87: 412-417.
Theodorou ME, Elrifi IR, Turpin DH, Plaxton WC (1991) Effects of phosphorus limitation on respiratory metabolism in the green alga Selenastrum minutum. Plant Physiology 95: 1089-1095.
Urban L, Le roux X, Sinoquet H, Jaffuel S, Jannoyer M (2003) A biochemical model of photosynthesis for mango leaves: evidence for the effect of fruit on photosynthetic capacity of nearby leaves. Tree Physiology 23: 289-300
Vieira G, Luiz FF, Luiz AE (1996) Growth and development of litchi cv. 'BrewsterC Bragantia 55: 325-328.
Wilkie JD, Sedgley M, Olesen T (2008) Regulation of floral initiation in horticultural trees. Journal of Experimental Botany 59: 3215-3228
Ying Z, Davenport T L (2004) Leaves Required for Floral Induction of Lychee. Plant Growth Regulation Society of America 32: 132-137