Artículo científico
La prohexadiona-Ca provoca cambios en el crecimiento vegetativo, giberelinas, rendimiento y luteolina en chile jalapeño
The prohexadione-Ca causes changes in vegetative growth, gibberellins, performance and luteolin in jalapeño pepper
1*Homero Ramírez, 1Víctor Manuel Camacho-Chávez, 1Lino Jeremías Ramírez-Pérez, 1José Hugo Rancaño-Arrioja, 2Leonardo Sepúlveda-Torre, 1Valentín Robledo-Torres
1Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo, 25315, Coah., México.
2Departamento de Investigación en Alimentos. Facultad de Ciencias Químicas. Universidad Autónoma de Coahuila. Blvd. Venustiano Carranza y José Cárdenas Col. República Ote. Saltillo, 25280, Coah., México.
*homeror@terra.com.mx
Recibido el 5 de junio de 2013
Aceptado el 12 de septiembre de 2014
RESUMEN.
En México, el chile jalapeño además de su aportación hortícola, requiere de nuevas tecnologías para mejorar su producción. La Prohexadiona de Calcio (P-Ca) es un retardante de crecimiento que mejora el rendimiento y la calidad del tomate. En esta investigación, se evaluó el efecto de la P-Ca en chile jalapeño híbrido Orozco a dosis de 0 (control-agua), 100, 125 y 175 mg L-1 aplicadas cuando la planta alcanzó 10 hojas verdaderas. Se evaluaron cada semana la altura y el diámetro de tallo. Además, se determinó el número de frutos y el rendimiento por planta. Se identificaron giberelinas por medio de cromatografía de gases-espectrometría de masas (CG-EM) en ápices muestreados 4 d después del tratamiento con P-Ca a 175 mg L-1 y se cuantificó por cromatografía liquida de alta resolución (CLAR) el contenido de luteolina en frutos maduros. Se observó una reducción de crecimiento del 30 % en la altura del tallo con P-Ca, este efecto desapareció al final del ciclo. El diámetro de tallo aumentó 20 % con P-Ca a 175 mg L-1. El rendimiento por planta se incrementó 15 % con 125 mg L-1 de P-Ca en relación al control. El número de frutos por planta no fue modificado. En los ápices del control, se identificaron las giberelinas A1, A4 y A7; mientras que en los tratados con P-Ca fueron detectadas A9, A20 y A51. El contenido de luteolina aumentó en frutos siete veces el nivel del testigo con P-Ca a 125 mg L-1.
Palabras clave: Antioxidante, Capsicum annuum L., hormonas, retardante de crecimiento.
ABSTRACT.
The cultivation of jalapeño chili in México, although makes an important and permanent horticultura! contribution, it requires a constantly adaptation of new technologies. Prohexadione-calcium (P-Ca) is a growth retardant which improves yield and fruit quality in tomato. In this study, it was evaluated the effect of P-Ca on jalapeño chili hybrid Orozco. The treatments consisted of: control (water), 100, 125 and 175 mg L-1 of P-Ca. They were applied when the plants reached 10 true leaves. The growth rate for height and diameter of main stem was weekly measured. Also, the number of fruits and yield per plant was determined. The identification of endogenous gibberellins at the apex from plants treated with P-Ca at 175 mg L-1 was conducted four days after treatment by gas chromatographymass spectrometry (GC-MS); whereas luteolin content was detected via HPLC in ripen fruits. The results indícate a reduction of 30 % in the rate of growth in stem height on treatments with P-Ca. This effect disappeared at the final stage of the growing process. Likewise, stem diameter increased 20 % with P-Ca at 175 mg L-1. The application of P-Ca at 125 mg L-1 resulted in a 15 % increase in yield when compared to the control. Fruit number was not affected. The gibberellins A1, A4 and A7 were found in control ápices; while in P-Ca samples A9, A20 and A51 were detected. Luteolin content in fruit was significantly increased x seven of control with P-Ca at 125 mg L-1.
Key words: Antioxidant, Capsicum annuum L., growth retardant, hormones.
INTRODUCCIÓN
El chile jalapeño es una hortaliza importante en la dieta de los habitantes y en la economía del país, ya que genera ingresos competitivos para los productores y favorece la creación de empleos (Macías-Duarte et al. 2012). Además, es una especie que requiere la constante aplicación de nuevas tecnologías en su manejo, con el propósito de que sea competitivo en el mercado nacional e internacional (Ramírez et al. 2009). Su fruto se caracteriza por la presencia de capsaicinoides y vitamina C (Vázquez-Flota et al. 2007). En años recientes se investiga la presencia de otros compuestos con valor fisiológico, donde destacan los flavonoides como la quercetina y la luteolina; compuestos que contribuyen a proteger el ADN de los radicales libres (Rosa et al. 2002, Díaz et al. 2004). La luteolina es un importante miembro del grupo de los flavonoides (Hernández et al. 2002). Los estudios realizados por Samejima et al. (1995) muestran un efecto quimioprotector de la luteolina contra tumores malignos in vivo sin efectos tóxicos colaterales; así como, propiedades antiinflamatorias. Este metabolíto actúa como antioxidante debido a su alta eficiencia en atrapar radicales libres y protege a las células del estrés oxidativo (Molina-Quijada et al. 2010).
Los biorreguladores son alternativas utilizadas en la agricultura contemporánea para modificar de forma temporal la acción de genes en plantas; lo que permite generar productos hortícolas que reúnan las características que demande el mercado (Rademacher 2000). La gran mayoría de los biorreguladores empleados en la producción de alimentos son amigables con el medio ambiente (Ramírez-Rodríguez et al. 2003). La prohexadiona de calcio (P-Ca) es un retardante del crecimiento prometedor para la horticultura moderna, la cual ha mostrado mejorar substancialmente la producción y calidad de frutos en diversas especies frutales caducifolias (Costa et al. 2004, Rademacher >et al. 2006). Inhibe la biosíntesis de las giberelinas A1, A4 y A7 biológicamente activas al bloquear la biosíntesis del 2-oxoglutarato dependiente de las dioxigenasas (Rademacher et al. 2006); una de las enzimas responsables de la biosíntesis de los flavonoides (Rademacher 2000). La presencia de P-Ca modifica la estructura y estimula la producción de flavonoides en hojas de manzano y peral (Prior y Cao 2000, Roemmelt et al. 2003a, Tosun y Ustun 2003). La investigación de la modificación o alteración de los niveles de antioxidantes por prácticas hortícolas es escasa. Al respecto Asa mi et al.(2003) y Kalt y Kushad (2000) reportaron que la investigación sobre el incremento en los niveles de antioxidantes en frutas y hortalizas es un tema de interés y representa una oportunidad en la estrategia actual dirigida al mejoramiento de la calidad de productos hortícolas. Por lo anterior, el objetivo del presente trabajo fue el evaluar el efecto de prohexadiona de calcio sobre el crecimiento, rendimiento, perfil de giberelinas endógenas en el ápice del tallo y contenido de luteolina en frutos maduros de chile jalapeño híbrido Orozco.
MATERIALES Y MÉTODOS
La presente investigación se llevó a cabo en el 2011 en la Universidad Autónoma Agraria Antonio Narro, Buenavista, Saltillo, Coahuila, México, ubicada en 25o 23' LN y 101o 01' LO, con altitud d de 1743 msnm. Se utilizó semilla de chile jalapeño hibrido Orozco, las cualse se sembraron el 1 de mayo en charolas de poliestireno de 200 cavidades, utilizando como sustrato Peat Moss Premier Mix. Cuando presentaron una altura promedio de 15 cm se trasplantaron de forma individual en bolsas de polietileno de 20 L que se llenaron con suelo de bosque. Las plantas se pusieron en un invernadero tipo cenital de estructura metálica con láminas laterales de policarbonato y plástico blanco lechoso (calibre 720) en el techo. Las cuales se ordenaron a distancia de 35 cm entre plantas y 75 cm entre hileras. Para la evaluación se utilizó un diseño experimental completamente al azar con 10 repeticiones. Cuando las plantas alcanzaron 10 hojas verdaderas, se aplicaron con un atomizador manual los siguientes tratamientos: control (agua), 100, 125 y 175 mg L-i de P-Ca, cada tratamiento se combinó con el surfactante líquido éternonifenol-polietilenglicol a dosis de 1 mL L-1. La aplicación se realizó entre las 8:00 y 9:00 am sobre el follaje de las plantas a punto de goteo. Las variables evaluadas fueron: altura y diámetro de tallo, número de frutos y rendimiento por planta; perfil de giberelinas en ápices por CG-EM (Ramírez et al. 2008) y cuantificación de luteolina en frutos maduros vía cromatografía liquida de alta resolución (CLAR) por medio de la técnica reportada por Samejima et al. (1995).
Altura y diámetro de tallo
La altura y diámetro de tallo se obtuvo al medirse cada semana durante todo el ciclo del cultivo. Para la altura, se utilizó una cinta métrica graduada en centímetros, midiendo desde la base del tallo hasta el ápice de la planta. Mientras que el diámetro de tallo se midió con un vernier en mm.
Número de frutos y rendimiento
El número de frutos y el rendimiento se determinaron al sumar los frutos cosechados y el peso de los mismos en cada uno de los cinco cortes realizados. Para pesar los frutos se utilizó una báscula Ohaus modelo 3729 con capacidad máxima de 3000 g y resolución de 0.1 g.
Análisis de giberelinas endógenas
Se utilizó una muestra de 1 g de peso seco; que se colocó en un matraz Erlenmeyer, al que se le agregaron 50 mL de metanol-agua (80:20 v/v) y se conservó durante 24 h en congelación (-15 °C). Posteriormente, se filtró en papel Wathman N°1 a temperatura de 24 °C. Esta actividad se repitió con el filtrado en dos ocasiones con igual cantidad de metanol (100 %) cada 4 h a la misma temperatura. Se reunieron los tres filtrados y se le eliminó el disolvente a presión reducida mediante un rotavaporador. Se procedió a la purificación preliminar de las giberelinas utilizando la técnica reportada por Ramírez et al. (2008) empleando cápsulas de Sep Pack C18. Enseguida los componentes de las muestras se separaron por cromatografía en capa fina preparativa (CCF) utilizando gel de sílice GF254, como eluyente se empleó el sistema isopropanol-amoniaco-agua (10:1:1) (v/v/v). Se visualizaron y compararon en luz ultravioleta con los estándares de las giberelinas A1, A4 y A7 localizadas en los Rf's (distancia que recorre la muestra en el plato de cromatografía de capa de fina con referencia a una giberelina conocida en un solvente específico) 0.4 a 0.7 respectivamente. Las giberelinas en los Rf's de cada muestra, fueron separadas y acondicionadas para su medición analítica (Stephan 1998). Cada muestra purificada fue metilada con diazometano preparado in situ, cada componente de la muestra purificada se inyectó (0.1 mL) en un cromatógrafo líquido de alta precisión modelo Finnigan TSQ 7000 equipado con nitrógeno como gas y una columna Ultrasep Es 100 RP18 de 1 m de largo por 0.43 mm de diámetro interno y empacada con acetonitrilo-agua conteniendo 0.2 % de ácido acético en proporción 50:50 (v/v) con un flujo de 70 mL min1. Las giberelinas A1, A4 y A7 fueron utilizadas como referencia analítica durante el programa de corridas por determinación cuantitativa de las giberelinas presentes en el tejido estudiado a través de la generación de la curva de calibración correspondiente, la cuál se construyó con 0.1 mL a concentraciones de 1, 10 y 100 ng de cada una de ellas diluidos en acetonametanol (50:50) (v/v).
Identificación de giberelinas por CG-EM
Las muestras con mayor actividad giberélica detectadas en la cromatografía de capa fina (CCF) fueron acondicionadas para identificar el tipo de giberelina con la técnica de cromatografía de gases y espectrometría de masas (Ramírez et ai. 2010). Se utilizó 1 g de peso seco por muestra referida la cual fue disuelta en 0.1 mL de acetona metanol (98 %) en la proporción 50:50 (v/v) y metilado con diazometano. Para la inyección al CG se prepararon previamente los derivados sililados. Una proporción del extracto metilado fue disuelto en 0.1 mL de piridina y tratado con 0.1 mL de trimetil clorosilano y hexametildisilazano. Las alícuotas fueron examinadas con un separador de membrana de Silicon Pye 104 CLC acoplado a un espectrómetro de masas AEI MS30. En este equipo se instalaron columnas de vidrio salinizadas (213 x 0.2 cm) con 2 % de Se-33, en 88-100 de gas chorm Q. La velocidad de flujo fue de 25 mL min-1 y la temperatura de la columna fue programada entre 180 y 280 °C a2 °C min-1. La espectrometría de masas fue determinada a 24 eV en una fuente de temperatura de 190 °C y una velocidad de búsqueda de 6.5 s por década de masas. El espectro fue registrado por una computadora Dec. Lin. 8. La identificación de giberelinas fue conducida por la comparación del índice de retención kovats (IRK) y el patrón de fragmentación de su espectrometría de masas de sus metil ester trimetil-silil éter con sus derivados de las muestras originales.
Extracción y cuantificación de luteolina por CLAR
El análisis de luteolina en frutos cosechados se realizó con la técnica reportada por Hernández et al. (2002). El material vegetal de cada muestra previamente congelado, se molió en un mortero aplicando de forma constante nitrógeno líquido. Luego se liofilizó y se colocaron 0.05 g en un tubo Ependorf de 1.5 mL al que se le agregó 1 mL de solución compuesta de metanol ácido fórmico agua destilada (70:1:29 v/v/v). La mezcla se sometió a vibración por 20 s en un agitador vortex, para luego sonicarlo por 40 min en un sonicador Bransom modelo 1510, para posteriormente centrifugarlo a 13,000 rpm en una microcentrífuga marca Labnet International modelo 24D por 10 min. Después de este proceso se recuperó el sobrenadante traslúcido con una jeringa. Esta solución se pasó a través de filtros siringe-driven filter unit de 0.45 µm (Millipore) y se colocó en un tubo Ependorf nuevo y etiquetado. Se desgasificó durante 5 min en un desgasificador Bransom modelo 1510 se inyectó en el cromatógrafo líquido de alta precisión marca Agilent modelo 1120, bajo el programa computacional EZChrom Elite Compact versión 33.0B. Las condiciones de corrida fueron: Columna Pursuit XRs, C-18, 150 x 4.6 mm, temperatura a 30 °C, fases: (A) metanol al 80 %, (B) acetonitrilo al 17 % y (C) ácido acético al 3 %, flujo de 1.0 mL min-1, volumen de inyección de 10 µL, longitud de onda de 254 nm y un tiempo de corrida de 30 min. La cantidad de luteolina en cada muestra se determinó con curvas estándar de calibración construidas previamente con luteolina (Sigma) en un rango de concentraciones de 0-500 -1.
Análisis estadísticos de los datos
Los resultados obtenidos se analizaron con el programa estadístico SAS (2009), para Windows versión 9.0 para obtener el análisis de varianza y la comparación de medias por medio de la prueba de Tukey ≥ 0.05.
RESULTADOS
Altura y diámetro de tallo
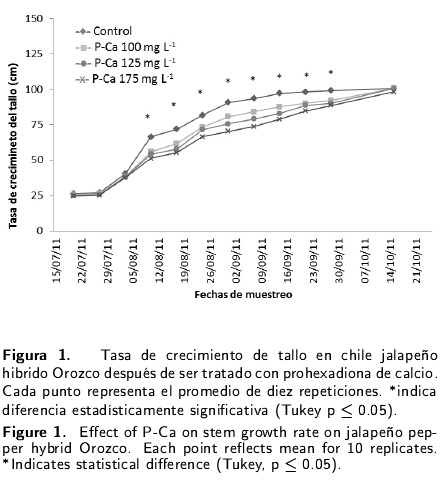
La prohexadiona de calcio modificó de forma significativa la tasa de crecimiento (P < 0.05) del tallo de plantas de chile jalapeño híbrido Orozco. Este efecto se observó a partir de la tercera semana de evaluación en los tratamientos con P-Ca (Figura 1). Las plantas tratadas con el retardante de crecimiento mostraron una menor altura durante la mayor parte de su ciclo fenológico. La menor reducción al compararse con el control, se reflejó entre 5 y 6 semanas después de la aplicación del retardante; siendo esta, de un 30 % en la dosis de 175 mg L-1 y de un 20 % en la dosis de 100 y 125 mg L-1. Este efecto desapareció al final de su ontogenia, cuando el P-Ca alcanzó al control. La tasa de crecimiento en el diámetro del tallo mostró un mayor desarrollo en los tratamientos con la P-Ca (Figura 2); sin embargo, la dosis de 175 mg L-1fue la que creció un 20 % más que el control. Lo que ocurrió a partir de la quinta semana después del tratamiento con la hormona.
Número de frutos y rendimiento
El rendimiento por planta fue estadísticamente mayor con la P-Ca a la dosis de 125 mg L-1 concentraciones del retardante a 100 y 175 mg L-1mostraron una tendencia a mayor y menor producción al compararse con el control (Tabla 1). El número de frutos por planta mostró una tendencia mayor en los tratamientos con la P-Ca; sin embargo, esta no fue significativa (Tabla 1).
Perfil de giberelinas endógenas
El análisis de cromatografía de gases y espectrometría de masas en ápices de chile jalapeño tratados con 175 mg L-1 de P-Ca, permitió identificar las giberelinas A9, A20 y A51. Mientras que en los ápices del testigo se encontraron las giberelinas A1, A4 y A7 (Tabla 2).
Contenido de luteolina
La aplicación de prohexadiona de calcio a cualquier dosis provocó un incremento de forma significativa (p > 0.05) en el contenido de luteolina en los frutos maduros (Figura 3)
El mayor aumento de este compuesto ocurrió en la dosis de 125 mg L-1, la cual superó siete veces al control; mientras que las dosis de 100 y 175 mg L-1 lo superaron tres y cuatro veces, respectivamente.
DISCUSIÓN
La reducción de crecimiento en el tallo de las plantas, fue evidente con la aplicación de la P-Ca. Este efecto también fue reportado por Altintas (2011) en plantas de tomate. En el presente estudio, las plantas tratadas con la hormona reanudaron su crecimiento durante las tres últimas semanas de su ciclo de desarrollo, que fue similar al del control al final del ciclo fenológico. Esto también ha sido observado en manzano (Ramírez-Rodríguez et al. 2003), chile mirador (Ramírez et al. 2010), uva (Lo Giudice et al. 2003, Lo Giudice et al. 2004), pera (Costa et al. 2004) y repollo (Ilias et al. 2007, Ouzounidou et al. 2010). El crecimiento del tallo está directamente ligado a la presencia y acción en ápices de las giberelinas biológicamente activas A1, A4, y A7 (Ramírez et al. 2010). La reducción en la tasa de crecimiento observada en este estudio (Figura 1), se atribuye al efecto de la P-Ca al inhibir la síntesis de estas giberelinas (Tabla 2). Las hormonas referidas son sintetizadas en los ápices del tallo en donde estimulan la división y la elongación celular; originando con ello, el crecimiento del tallo; sin embargo, la presencia de la P-Ca en el tejido apical, inhibe este proceso fisiológico al bloquear temporalmente la síntesis de estas giberelinas (Rademacher 2000). Durante la ausencia de esas hormonas, las giberelinas biológicamente inactivas A9, A20 y A51 estuvieron presentes. Este cambio en el perfil de giberelinas ha sido explicado por Rademacher y Kober (2003) a través del mecanismo que ejerce la P-Ca al inhibir la 3β-hidroxilación de las hormonas AG9y AG20 a AG1 y AG4 por las AG3 oxidasas. Lo anterior, se confirma con los reportes en manzano (Rademacher 2004), tomate y chile pimiento (Ramírez et al. 2008). Al final de la fase de desarrollo, las plantas tratadas con la P-Ca restauraron su crecimiento. Lo anterior, puede reflejar la recuperación del sistema metabólico en la síntesis de las giberelinas Ai, A4, y A7. Estos efectos son una característica observada en durazno y pera (Byers et al. 2004). En estas especies se demostró que el bloqueo en la síntesis de giberelinas activas por la P-Ca es temporal, ya que las AG1, AG4 y AG7 son nuevamente detectadas previo a la reactivación del crecimiento (Rademacher 2004). El incremento en el diámetro del tallo ha sido también observado en otros cultivos hortícolas (Ramírez et al. 2005). En papa se observó un efecto similar cuando se aplicó el retardante paclobutrazol observándose un aumento en la síntesis de almidón en las células de médula y corticales del tallo (Tsegaw et al. 2005) y una acumulación de otros asimilados al reducirse la demanda de ellos en el meristemo apical (Ramírez-Rodríguez et al. 2003). En este estudio, es probable que la P-Ca haya actuado en una forma similar; por lo que sería un tema interesante de una investigación futura.
El aumento en el rendimiento cuando la P-Ca fue aplicada en el rango del presente estudio ha sido reportado en tomate (Ramírez et al. 2005, Gemici et al. 2007, Ramírez et al. 2008) y chile (Ravi-raja et al. 2008, Sridhar et al. 2009, Ramírez et al. 2010). El aumento en el rendimiento observado con la aplicación de la P-Ca podría reflejar la influencia que tuvo el retardante de crecimiento en una mayor translocación de asimilados del ápice hacia los frutos presentes (Rademacher 2000). Lo anterior, causado a través de la ausencia de las giberelinas A1, A4 y A7 (Tabla 2). De esta manera, los frutos que permanecieron en la planta recibieron mayor flujo de alimento hasta el término de su ciclo (Costa et al. 2004, Rademacher 2004, Sridhar et al. 2009, Flores et al. 2011).
El incremento de luteolina observado en los frutos de plantas tratadas con cualquier dosis del retardante es notable al compararse con el control (Figura 3). El mayor contenido de este compuesto ocurrió con la concentración de la P-Ca a 125 mg L-1. Al compararlo con el menor efecto observado en las dosis a 100 y 175 mg L-i se refleja la propuesta de Ramírez et al. (2008) en términos de que la hormona actuó al nivel subóptimo y supraóptimo en el proceso de producción de luteolina. La prohexadiona de calcio es un retardante de crecimiento que al ser aplicado en hortalizas como tomate influye en la biosíntesis de los flavonoides como las 3-desoxicatequinas (Rademacher 2000). Lo anterior ocurre por el efecto que tiene la molécula de P-Ca sobre el 2-oxoglutarato que produce dioxigenasas (Roemmelt et al. 1999). Al respecto Roemmelt et al. (2003b) encontraron en manzano y peral que en aplicaciones entre 30 y 250 mg L-1 que la P-Ca actúa sobre la enzima hidrolasaflavona (FHT), originando un bloqueo en la síntesis de quercetina y un estímulo en la formación de la enzima flavona 4 reductasa, activa en la síntesis del luteoferol, que promueve la formación de luteolina. Mientras que Rademacher et al. (2006) sugieren que la P-Ca estimula la síntesis de eriodictol-7-glucosido que es un precursor del luteoferol, estableciendo correlación entre el contenido de luteolina y procianidina. Sobre lo mismo, Treutter et al. (2010) reportaron en actinidia un aumento en los niveles de flavonoides cuando aplicaron P-Ca combinado con citocininas y giberelinas. Los reportes y los resultados de la Figura 3 permiten considerar un indicio del sinergismo que existe entre la P-Ca y las hormonas endógenas de la planta sobre la biosíntesis de algunos flavonoides. Este efecto a su vez, puede provocar cambios positivos en las plantas hortícolas. Lo anterior, ha sido demostrado con mayor floración y aumento de rendimiento en guayaba (Vargas-Álvarez et al. 2005), manzano (Ramírez-Rodríguez et al. 2003) y chile (Ramírez et al. 2010). El mecanismo de acción protectora de la luteolina en plantas no se ha elucidado; sin embargo se ha propuesto que esta sustancia natural evita daños en el ADN, causados por radicales libres inducidos por factores adversos internos y externos. Esta condición define a la luteolina con mayor actividad antimutagénica al compararse con otros antioxidantes como la quercetina (Samejima et al. 1995, Razmara et al. 2010).
CONCLUSIONES
Se concluye que la aplicación de prohexadiona de calcio en chile jalapeño híbrido Orozco reduce de forma temporal y significativa la tasa de crecimiento de la altura en un 30 % y aumenta el diámetro del tallo en un 20 %; inhibe la síntesis de las giberelinas A1, A4 y A7 en el ápice, incrementa el rendimiento en un 15 % y el contenido de luteolina hasta siete veces en los frutos maduros.
AGRADECIMIENTOS
Los autores agradecen al B.S. Dan Flick (Wilburellis, USA) por haber proporcionado la prohexadiona de calcio utilizada en esta investigación.
LITERATURA CITADA
Altintas S (2011) Effects of prohexadione-calcium with three rates of phosphorus and chlormequat chloride on vegetative and generative growth of tomato. African Journal of Biotechnology 10: 17142-17151.
Asami KD, Hong YJ, Barrett DM, Mitchell AE (2003) Comparison of the total phenolic and ascorbic acid content of freeze-dried and air-dried marionberry, strawberry, and corn grown using conventional, organic, and sustainable agricultural practices. Journal of Agricultural and Food Chemistry 51: 1237-1241.
Byers RE, Carbaugh DH, Combs LD (2004) Prohexadione-calcium suppression of apple tree shoot growth as affected by spray additives. Hortscience 39: 115-119.
Costa G, Sabatini E, Spinelli F, Andreotti C, Bomben C, Vizzotto G (2004) Two years of application of prohexadione-Ca on apple: Effect on vegetative and cropping performance, fruit quality, return bloom and residual effect. Acta Horticulturae 653: 35-40.
Díaz J, Pomar F, Bernal A, Merino F (2004) Peroxidases and the metabolism of capsaicin in Capsicum annuum L. Phytochemistry Reviews 3: 141-157.
Flores LR, Sánchez DF, Rodríguez JE, Mora AR, Colinas LMT, Lozoya SH (2011) Paclobutrazol, uniconazol y cycocel en la producción de tubérculosemilla de papa en cultivo hidropónico. Revista Chapingo Serie Horticultura 17: 173-182.
Gemici M, Türkyilmaz B, Tan K (2007) Effects of 2, 4-D and 4-CPA on yield and quality of the tomato, Lycopersicon esculentum Miller (Solanaceae). Ege Universitesi. Fen Fakultesi Dergisi. Seri A 29: 24-32.
Hernández JD, García L, Hernández A, Álvarez R, Román LU (2002) Glicósidos de luteolina y miricetina de burseraceae. Revista de la Sociedad Química de México 46: 295-300.
Ilias I, Ouzounidou G, Giannakoula, Papadopoulou P (2007) Effects of gibberellic acid and prohexadione-calcium on growth, chlorophyll, fluorescence and quality of okra plant. Biología Plantarum 51: 575-578.
Kalt W, Kushad MM (2000) The role of oxidative stress and antioxidants in plants and human health: Introduction to the colloquium. Hortscience 35: 572-574.
Lo Giudice D, Wolf TK, Marini RP (2003) Vegetative response of Vitis vinifera to prohexadione-calcium. Hortscience 38: 1435-1438.
Lo Giudice D, Wolf TK, Zoeckelin BW (2004) Effects of prohexadione-calcium on grape yield components and fruit and wine composition. American Journal of Enology and Viticulture 55: 73-83.
Macías-Duarte R, Grijalva-Contreras RL, Robles-Contreras F (2012) Respuesta de la aplicación de estiércol y fertilizantes sobre el rendimiento y calidad de chile jalapeño. Revista de Ciencias Biológicas y de la Salud 14: 32-38.
Molina-Quijada DMA, Medina-Juárez LA, Gonzáles-Aguilar GA, Robles-Sánchez RM, Gámez-Meza N (2010) Phenolic compounds and antioxidant activity of table grape (Vitis vinifera L.) skin from northwest México. CyTA - Journal of Food 8: 57-63.
Ouzounidou G, Ilias L, Gianna-Koula A, Papadoupulou P (2010) Comparative study on the effects of various plant growth regulators on growth, quality and physiology of Capsicum annuum L. Pakistán Journal of Botany 42: 805-814.
Prior RL, Cao G (2000) Antioxidant phytochemicals in fruits and vegetables: Diet and health implications. Hortscience 35: 588-592.
Rademacher W (2000) Growth retardants: Effects on gibberellin biosynthesis and other metabolic pathways. Annual Review of Plant Physiology and Plant Molecular Biology 51: 501-531.
Rademacher W (2004) Chemical regulation of shoot growth in fruit trees. Acta Horticulturae 653: 29- 32.
Rademacher W, Kober L (2003) Efficient use of prohexadione-Ca in pome fruits. European Journal of Horticultural Science 68: 107-107.
Rademacher W, Spinelli F, Costa G (2006) Prohexadione Ca: Modes of action of a multifunctional plant bioregulator for fruit trees. Acta Horticuturae 727: 97-106.
Ramírez-Rodríguez H, Gómez-Castañeda JC, Benavides-Mendoza A, Robledo-Torres V, Encina-Rodríguez Ll, Coello-Cutiño CA (2003) Influencia de prohexadiona-Ca sobre crecimiento vegetativo, producción y calidad de fruto en manzano. Revista Chapingo Serie Horticultura 9: 279-284.
Ramírez H, Peralta-Manjarrez RM, Benavides-Mendoza A, Sánchez-López A, Robledo-Torres V, Hernández-Dávila J (2005) Efectos de prohexadiona-Ca en tomate y su relación con la variación de la concentración de giberelinas y citocininas. Revista Chapingo Serie Horticultura 11: 283-290.
Ramírez H, Herrera-Gámez B, Méndez-Quiroz YH, Benavides-Mendoza A, De La Cruz-Bretón JA, Álvarez-Mares V, Raneaño-Arrioja JH, Villareal-Quintanilla JA (2008) Prohexadiona de calcio disminuye el contenido de giberelinas endógenas en ápices de tomate saladette y chile pimiento. Revista Chapingo Serie Horticultura 14: 193-198.
Ramírez H, Méndez-Paredes O, Benavides-Mendoza A, Amado-Ramírez C (2009) Influencia de prohexadiona-Ca y promotores de oxidación sobre el rendimiento, capsaicina y vitamina C en chile jalapeño. Revista Chapingo Serie Horticultura 15: 231-236
Ramírez H, Amado-Ramírez C, Benavides-Mendoza A, Robledo-Torres V, Martínez-Osorio A (2010) Prohexa-diona-Ca, AG3, ANOXA y BA modifican indicadores fisiológicos en chile mirador. Revista Chapingo Serie Horticultura 16: 83-89.
Raviraja SG, Krishna MR, Vishwanath AP, Kempegowda K, Raghavendra D (2008) Influence of pruning and growth regulators on the shelf life of coloured capsicum (Capsicum annuum L.) cv. Bombi under greenhouse. Journal Agricultural Science 42: 33-37.
Razmara RS, Daneshfar A, Sahraei R (2010) Solubility of quercetin in water + methanol and water + ethanol from (292.8 to 333.8) K. Journal Chemical Engineering Data 55: 3934-3936.
Roemmelt S, Treutter D, Speakman JB, Rademach-Er W (1999) Effects of Prohexadione-Ca on the flavonoid metabolism of apple with respect to plant resistance against fire blight. Acta Horticulturae 489: 359364.
Roemmelt S, Zimmermann N, Rademacher W, Treutter D (2003a) Formation of novel flavonoids in apple (Malus x domestica) treated with the 2-oxoglutarate-dependent dioxygenase inhibitor Prohexadione-Ca. Phytochemistry 64: 709-716.
Roemmelt S, Fischer TC, Halbwirth H, Peterek S, Schlangen K, Speakman JB, Treutter D, Forkmann G, Stich K (2003b) Effect of dioxygenase inhibitors on the resistance-related flavonoid metabolism of apple and pears: Chemical, biochemical and molecular biological aspeets. European Journal Horticulture Science 68: 129-136.
Rosa A, Deiana M, Casu V, Paccagnini S, Appendino G, Ballero M, Assunta DM (2002) Antioxidant activity of capsinoids. Journal of Agricultural and Food Chemistry 50: 7396-7401.
SAS, Institute (2009) SAS/STAT. Users Guide. Release 9.00 ed. SAS Institute Inc., Cary, North Carolina., USA.
Samejima K, Kanazawa K, Ashida H, Danno G (1995) Luteolin: A strong antimutagen against dietary carcinogen, Trp-P-2, in peppermint, sage and thyme. Journal of Agricultural and Food Chemistry 43: 410-414.
Sridhar G, Koti RV, Chetti MB, Hiremath SM (2009) Effect of naphthalene acetic acid and mepiquat chloride on physiological components of yield in bell pepper (Capsicum annuum L.). Journal Agriculture Research 47: 53-62.
Stephan M, Bangerth F, Schneider G (1998) Transport and metabolism of the gibberellins A1, A3 and A4 after application to developing apple fruits of Malus domestica cv. Jonagold. Acta Florticulturae 463: 113-120.
Tosun I, Ustun NS (2003) An investigation about antioxidant capacity of fruit nectars. Pakistán Journal of Nutrition 2: 167-169.
Treutter D, Hadersdorfer J, Pietzner J, Steber M (2010) Effect of bioregulators on growth and secondary metabolism of Actinidia arguta plants. Acta Horticulturae 884: 689-694.
Tsegaw T, Hammes S, Robbertse J (2005) Paclobutrazolinduced leaf, stem, and root anatomical modification in potato. Hortscience 40: 1343-1346.
Vargas-Álvarez D, Soto-Hernández M, González-Hernández VA, Engleman EM, Martínez-Garza A (2005) Variación del contenido de flavonoides en hojas de guayaba en condiciones de estrés. Revista Chapingo Serie Horticultura 11: 89-92.
Vázquez-Flota F, Miranda-Ham ML, Monforte-González M, Gutiérrez-Carbajal G, Velázquez-García C, Nieto-Pelayo Y (2007) La biosíntesis de capsaicinoides, el principio picante del chile. Revista Fitotecnia Mexicana 30: 353-360.